Related Research Articles

Sleep is a naturally recurring state of mind and body, characterized by altered consciousness, relatively inhibited sensory activity, reduced muscle activity and inhibition of nearly all voluntary muscles during rapid eye movement (REM) sleep, and reduced interactions with surroundings. It is distinguished from wakefulness by a decreased ability to react to stimuli, but more reactive than a coma or disorders of consciousness, with sleep displaying different, active brain patterns.

A dream is a succession of images, ideas, emotions, and sensations that usually occur involuntarily in the mind during certain stages of sleep. During a typical lifespan, a person spends a total of about six years dreaming. Most dreams last only 5 to 20 minutes.
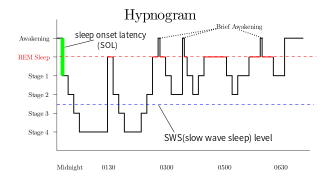
Rapid eye movement sleep is a unique phase of sleep in mammals and birds, characterized by random rapid movement of the eyes, accompanied by low muscle tone throughout the body, and the propensity of the sleeper to dream vividly.

The neocortex, also called the neopallium, isocortex, or the six-layered cortex, is a set of layers of the mammalian cerebral cortex involved in higher-order brain functions such as sensory perception, cognition, generation of motor commands, spatial reasoning and language. The neocortex is further subdivided into the true isocortex and the proisocortex.

The claustrum is a thin, bilateral collection of neurons and supporting glial cells, that connects to cortical and subcortical regions of the brain. It is located between the insula laterally and the putamen medially, separated by the extreme and external capsules respectively. The blood supply to the claustrum is fulfilled via the middle cerebral artery. It is considered to be the most densely connected structure in the brain, allowing for integration of various cortical inputs into one experience rather than singular events. The claustrum is difficult to study given the limited number of individuals with claustral lesions and the poor resolution of neuroimaging.
Non-rapid eye movement sleep (NREM), also known as quiescent sleep, is, collectively, sleep stages 1–3, previously known as stages 1–4. Rapid eye movement sleep (REM) is not included. There are distinct electroencephalographic and other characteristics seen in each stage. Unlike REM sleep, there is usually little or no eye movement during these stages. Dreaming occurs in both during both sleep states, and muscles are not paralyzed as in REM sleep. People who do not go through the sleeping stages properly get stuck in NREM sleep, and because muscles are not paralyzed a person may be able to sleepwalk. According to studies, the mental activity that takes place during NREM sleep is believed to be thought-like, whereas REM sleep includes hallucinatory and bizarre content. NREM sleep is characteristic of dreamer-initiated friendliness, compared to REM sleep where it's more aggressive, implying that NREM is in charge of simulating friendly interactions. The mental activity that occurs in NREM and REM sleep is a result of two different mind generators, which also explains the difference in mental activity. In addition, there is a parasympathetic dominance during NREM. The reported differences between the REM and NREM activity are believed to arise from differences in the memory stages that occur during the two types of sleep.

A K-complex is a waveform that may be seen on an electroencephalogram (EEG). It occurs during stage 2 of NREM sleep. It is the "largest event in healthy human EEG". They are more frequent in the first sleep cycles.
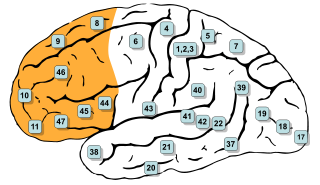
In mammalian brain anatomy, the prefrontal cortex (PFC) is the cerebral cortex which covers the front part of the frontal lobe. The PFC contains the Brodmann areas BA8, BA9, BA10, BA11, BA12, BA13, BA14, BA24, BA25, BA32, BA44, BA45, BA46, and BA47.

Neural oscillations, or brainwaves, are rhythmic or repetitive patterns of neural activity in the central nervous system. Neural tissue can generate oscillatory activity in many ways, driven either by mechanisms within individual neurons or by interactions between neurons. In individual neurons, oscillations can appear either as oscillations in membrane potential or as rhythmic patterns of action potentials, which then produce oscillatory activation of post-synaptic neurons. At the level of neural ensembles, synchronized activity of large numbers of neurons can give rise to macroscopic oscillations, which can be observed in an electroencephalogram. Oscillatory activity in groups of neurons generally arises from feedback connections between the neurons that result in the synchronization of their firing patterns. The interaction between neurons can give rise to oscillations at a different frequency than the firing frequency of individual neurons. A well-known example of macroscopic neural oscillations is alpha activity.

Neural binding is the neuroscientific aspect of what is commonly known as the binding problem: the interdisciplinary difficulty of creating a comprehensive and verifiable model for the unity of consciousness. "Binding" refers to the integration of highly diverse neural information in the forming of one's cohesive experience. The neural binding hypothesis states that neural signals are paired through synchronized oscillations of neuronal activity that combine and recombine to allow for a wide variety of responses to context-dependent stimuli. These dynamic neural networks are thought to account for the flexibility and nuanced response of the brain to various situations. The coupling of these networks is transient, on the order of milliseconds, and allows for rapid activity.
Recurrent thalamo-cortical resonance is an observed phenomenon of oscillatory neural activity between the thalamus and various cortical regions of the brain. It is proposed by Rodolfo Llinas and others as a theory for the integration of sensory information into the whole of perception in the brain. Thalamocortical oscillation is proposed to be a mechanism of synchronization between different cortical regions of the brain, a process known as temporal binding. This is possible through the existence of thalamocortical networks, groupings of thalamic and cortical cells that exhibit oscillatory properties.

The neural correlates of consciousness (NCC) constitute the minimal set of neuronal events and mechanisms sufficient for a specific conscious percept. Neuroscientists use empirical approaches to discover neural correlates of subjective phenomena; that is, neural changes which necessarily and regularly correlate with a specific experience. The set should be minimal because, under the assumption that the brain is sufficient to give rise to any given conscious experience, the question is which of its components is necessary to produce it.
Ponto-geniculo-occipital waves or PGO waves are distinctive wave forms of propagating activity between three key brain regions: the pons, lateral geniculate nucleus, and occipital lobe; specifically, they are phasic field potentials. These waves can be recorded from any of these three structures during and immediately before REM sleep. The waves begin as electrical pulses from the pons, then move to the lateral geniculate nucleus residing in the thalamus, and end in the primary visual cortex of the occipital lobe. The appearances of these waves are most prominent in the period right before REM sleep, albeit they have been recorded during wakefulness as well. They are theorized to be intricately involved with eye movement of both wake and sleep cycles in many different animals.
Memory consolidation is a category of processes that stabilize a memory trace after its initial acquisition. A memory trace is a change in the nervous system caused by memorizing something. Consolidation is distinguished into two specific processes. The first, synaptic consolidation, which is thought to correspond to late-phase long-term potentiation, occurs on a small scale in the synaptic connections and neural circuits within the first few hours after learning. The second process is systems consolidation, occurring on a much larger scale in the brain, rendering hippocampus-dependent memories independent of the hippocampus over a period of weeks to years. Recently, a third process has become the focus of research, reconsolidation, in which previously consolidated memories can be made labile again through reactivation of the memory trace.
Charcot–Wilbrand syndrome (CWS) describes dream loss following focal brain damage specifically characterised by visual agnosia and loss of ability to mentally recall or "revisualize" images. The name of this condition dates back to the case study work of Jean-Martin Charcot and Hermann Wilbrand, and was first described by Otto Potzl as “mind blindness with disturbance of optic imagination”. MacDonald Critchley, former president of the World Federation of Neurology, more recently summarized CWS as “a patient loses the power to conjure up visual images or memories, and furthermore, ceases to dream during his sleeping hours”. This condition is quite rare and affects only a handful of brain damage patients. Further study could help illuminate the neurological pathway for dream formation.

The relationship between sleep and memory has been studied since at least the early 19th century. Memory, the cognitive process of storing and retrieving past experiences, learning and recognition, is a product of brain plasticity, the structural changes within synapses that create associations between stimuli. Stimuli are encoded within milliseconds; however, the long-term maintenance of memories can take additional minutes, days, or even years to fully consolidate and become a stable memory that is accessible. Therefore, the formation of a specific memory occurs rapidly, but the evolution of a memory is often an ongoing process.
Secondary consciousness is an individual's accessibility to their history and plans. The ability allows its possessors to go beyond the limits of the remembered present of primary consciousness. Primary consciousness can be defined as simple awareness that includes perception and emotion. As such, it is ascribed to most animals. By contrast, secondary consciousness depends on and includes such features as self-reflective awareness, abstract thinking, volition and metacognition. The term was coined by Gerald Edelman.
The activation-synthesis hypothesis, proposed by Harvard University psychiatrists John Allan Hobson and Robert McCarley, is a neurobiological theory of dreams first published in the American Journal of Psychiatry in December 1977. The differences in neuronal activity of the brainstem during waking and REM sleep were observed, and the hypothesis proposes that dreams result from brain activation during REM sleep. Since then, the hypothesis has undergone an evolution as technology and experimental equipment has become more precise. Currently, a three-dimensional model called AIM Model, described below, is used to determine the different states of the brain over the course of the day and night. The AIM Model introduces a new hypothesis that primary consciousness is an important building block on which secondary consciousness is constructed.

The neuroscience of sleep is the study of the neuroscientific and physiological basis of the nature of sleep and its functions. Traditionally, sleep has been studied as part of psychology and medicine. The study of sleep from a neuroscience perspective grew to prominence with advances in technology and proliferation of neuroscience research from the second half of the twentieth century.

Prefrontal synthesis is the conscious purposeful process of synthesizing novel mental images. PFS is neurologically different from the other types of imagination, such as simple memory recall and dreaming. Unlike dreaming, which is spontaneous and not controlled by the prefrontal cortex (PFC), PFS is controlled by and completely dependent on the intact lateral prefrontal cortex. Unlike simple memory recall that involves activation of a single neuronal ensemble (NE) encoded at some point in the past, PFS involves active combination of two or more object-encoding neuronal ensembles (objectNE). The mechanism of PFS is hypothesized to involve synchronization of several independent objectNEs. When objectNEs fire out-of-sync, the objects are perceived one at a time. However, once those objectNEs are time-shifted by the lateral PFC to fire in-phase with each other, they are consciously experienced as one unified object or scene.
References
- ↑ Shepherd, Michael (1983). "Sleeping and dreaming". British Medical Journal (Clinical Research Ed.). 287 (6391): 512–513. doi:10.1136/bmj.287.6391.512. PMC 1548456 . PMID 6411198.
- ↑ Crick, Francis; Mitchison, Graeme (1983). "The function of dream sleep". Nature. 304 (5922): 111–4. Bibcode:1983Natur.304..111C. doi:10.1038/304111a0. PMID 6866101. S2CID 41500914.
- ↑ Hopfield, John J.; Feinstein, David I.; Palmer, Richard G. (14 July 1983). " 'Unlearning' has a stabilizing effect in collective memories". Nature. 304 (5922): 158–159. Bibcode:1983Natur.304..158H. doi:10.1038/304158a0. PMID 6866109. S2CID 4269710.
- ↑ Hobson; McCarley, RW (1977). "The brain as a dream state generator: an activation-synthesis hypothesis of the dream process". The American Journal of Psychiatry. 134 (12): 1335–48. doi:10.1176/ajp.134.12.1335. PMID 21570.
- ↑ Crick, Francis (1988). "Neural networks and REM sleep". Bioscience Reports. 8 (6): 531–5. doi:10.1007/BF01117331. PMID 3242640. S2CID 31962759.
- ↑ Allison, T; Cicchetti, D. (1976). "Sleep in mammals: ecological and constitutional correlates". Science. 194 (4266): 732–4. Bibcode:1976Sci...194..732A. doi:10.1126/science.982039. PMID 982039.
- ↑ Crick; Mitchison, G (1983). "The function of dream sleep". Nature. 304 (5922): 111–4. Bibcode:1983Natur.304..111C. doi:10.1038/304111a0. PMID 6866101. S2CID 41500914.
- ↑ Osame Kinouchi; Renato Rodrigues Kinouchi (2002). "Dreams, endocannabinoids and itinerant dynamics in neural networks: re elaborating Crick-Mitchison unlearning hypothesis". arXiv: cond-mat/0208590v2 .