
Archosauromorpha is a clade of diapsid reptiles containing all reptiles more closely related to archosaurs rather than lepidosaurs. Archosauromorphs first appeared during the late Middle Permian or Late Permian, though they became much more common and diverse during the Triassic period.

Hyperodapedon is an extinct genus of rhynchosaur reptiles which lived during Late Triassic period. Like other rhynchosaurs, it was an heavily built archosauromorph, distantly related to archosaurs such as crocodilians and dinosaurs. Hyperodapedon in particular was part of the subfamily Hyperodapedontinae, a specialized rhynchosaurian subgroup with broad skulls, beaked snouts, and crushing tooth plates on the roof of the mouth.

The Ischigualasto Formation is a Late Triassic geological formation in the Ischigualasto-Villa Unión Basin of southwestern La Rioja Province and northeastern San Juan Province in northwestern Argentina. The formation dates to the late Carnian and early Norian stages of the Late Triassic, according to radiometric dating of ash beds.
Mesosuchus is an extinct genus of basal Rhynchosaur from early Middle Triassic deposits of Eastern Cape, South Africa. It is known from the holotype SAM 5882, a partial skeleton, and from the paratypes SAM 6046, SAM 6536, SAM 7416 and SAM 7701 from the Aliwal North Euparkeria site. Mesosuchus is quite small, spanning around 30 cm in length. Mesosuchus was discovered and named by David Meredith Seares Watson in 1912.

Proterochampsa is an extinct genus of proterochampsid archosauriform from the Late Triassic of South America. The genus is the namesake of the family Proterochampsidae, and the broader clade Proterochampsia. Like other proterochampsids, Proterochampsa are quadruped tetrapods superficially similar in appearance to modern crocodiles, although the two groups are not closely related. Proterochampsids can be distinguished from other related archosauriformes by characters such as a dorsoventrally flattened, triangular skull with a long, narrow snout at the anterior end and that expands transversally at the posterior end, asymmetric feet, and a lack of postfrontal bones in the skull, with the nares located near the midline. Proterochampsa is additionally defined by characters of dermal sculpturing consisting of nodular protuberances on the skull, antorbital fenestrae facing dorsally, and a restricted antorbital fossa on the maxilla. The genus comprises two known species: Proterochampsa barrionuevoi and Proterochampsa nodosa. P. barrionuevoi specimens have been discovered in the Ischigualasto Formation in northwestern Argentina, while P. nodosa specimens have been found in the Santa Maria supersequence in southeastern Brazil. The two species are distinct in several characters, including that P. nodosa has larger, more well-developed nodular protuberances, a more gradually narrowing snout, and a higher occiput than P. barrionuevoi. Of the two, P. nodosa is thought to have less derived features than P. barrionuevoi.

Bentonyx is an extinct genus of rhynchosaur from the middle Triassic epoch of Devon in England. Its fossil, a well preserved skull, BRSUG 27200, was discovered in Otter Sandstone Formation and was first assigned to Rhynchosaurus spenceri, that is known from 25 specimens. This species was reassigned to its own genus, Fodonyx, that was first described by David W. E. Hone and Michael Benton in 2008. More recently, this skull was reassigned to this genus by Max C. Langer, Felipe C. Montefeltro, David E. Hone, Robin Whatley and Cesar L. Schultz in 2010 and the type species is Bentonyx sidensis.

Fodonyx is an extinct genus of rhynchosaur from the middle Triassic epoch of Devon in England. Its fossils were discovered in Otter Sandstone Formation and were first assigned to Rhynchosaurus spenceri. This species was reassigned to its own genus, Fodonyx the holotype of which is EXEMS 60/1985/292, that described by David W. E. Hone and Michael J. Benton in 2008. More recently, one skull was reassigned to the new genus Bentonyx. It is distinguished from other rhynchosaurs by a single autapomorphy, the ventral angling of the paraoccipital processes. In all other rhynchosaurs these processes angle dorsally or are horizontal. It is not known if this conferred any advantage to Fodonyx. Fodonyx was between 40 and 50 cm long.
Teyumbaita is an extinct genus of hyperodapedontine rhynchosaur from the Upper Triassic of southern Brazil. Its fossils were recovered from the early Norian-age Caturrita Formation, one of several fossiliferous formations exposed at Paleorrota Geopark in the state of Rio Grande do Sul. Teyumbaita is likely the youngest valid genus of rhynchosaur, as other members of the group likely died out before the start of the Norian.

Stenaulorhynchus is an extinct genus of hyperodapedontid rhynchosaur known from the Middle Triassic deposits of Tanganyika Territory, Tanzania. It was found in the Lifua Member of the Manda Formation in the Karoo Supergroup. It was named and first described by Sidney Henry Haughton in 1932. The type species is Stenaulorhynchus stockleyi, a beaked herbivore measuring 1–6 meters in length.
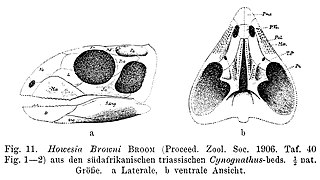
Howesia is an extinct genus of basal rhynchosaur from early Middle Triassic deposits of Eastern Cape, South Africa. It is known from the holotype SAM 5884, a partial skeleton with palate and partial lower jaws and from two paratypes, SAM 5885 and SAM 5886. It was found in the Burgersdorp Formation of the middle deposits of the Beaufort Group and referred to Subzone B of the Cynognathus Assemblage Zone. It was first named by Robert Broom in 1905 and the type species is Howesia browni, named after Alfred Brown.

Hyperodapedontinae is a subfamily of rhynchosaurs within the family Rhynchosauridae. Fossils have been found from Argentina, Brazil, Canada, India, Madagascar, Scotland, Tanzania, United States and Zimbabwe.
Macrocephalosaurus is a genus of rhynchosaurs from the Late Triassic period of southern Brazil. It contains a single species, Macrocephalosaurus mariensis. Although usually synonymized with Hyperodapedon, some cladistic analyses have called this synonymy into question.
Otischalkia is an extinct genus of archosauromoph from late Triassic deposits of Howard County, Texas, US It is known from the holotype TMM 31025-263, left humerus and from the referred specimens TMM 31025-262, TMM 31025-266, TMM 31025-264, TMM 31185-92 and TMM 31185-93. It was found in the Lower Dockum Group near the abandoned settlement of Otis Chalk. It was first named by Adrian P. Hunt and Spencer G. Lucas in 1991 and the type species is Otischalkia elderae.

Teraterpeton is an extinct genus of trilophosaurid archosauromorphs. It is known from a partial skeleton from the Late Triassic Wolfville Formation of Nova Scotia, described in 2003. It has many unique features seen in no other related form, including an elongated, toothless snout and large openings for the nostrils. Because of this, Teraterpeton was originally placed in its own family, Teraterpetidae, related to Trilophosaurus. Newer studies generally place it within Trilophosauridae.

Prolacerta is a genus of archosauromorph from the lower Triassic of South Africa and Antarctica. The only known species is Prolacerta broomi. The generic name Prolacerta is derived from Latin meaning “before lizard” and its species name broomi is in commemoration of the famous paleontologist Robert Broom, who discovered and studied many of the fossils found in rocks of the Karoo Supergroup. When first discovered, Prolacerta was considered to be ancestral to modern lizards, scientifically known as lacertilians. However, a study by Gow (1975) instead found that it shared more similarities with the lineage that would lead to archosaurs such as crocodilians and dinosaurs. Prolacerta is considered by modern paleontologists to be among the closest relatives of the Archosauriformes.

Noteosuchus is an extinct genus of basal rhynchosaur known from the earliest Triassic deposits of Eastern Cape Province, South Africa. It was first named by David Meredith Seares Watson in 1912 and the type species is Eosuchus colletti. The generic name Eosuchus is preoccupied by the generic name of Eosuchus lerichei Dollo, 1907, a gavialoid crocodilian known from northern France. Thus, an alternative generic name, Noteosuchus, was proposed by Robert Broom in 1925. The generic name erected by Broom (1925) is a compound, meaning "Not Eosuchus", while "Eosuchus" is derived from the name of Eos, the goddess of the dawn in Greek mythology, and suchus, Latinized from the Greek souchos, an Egyptian crocodile god, thus meaning "dawn crocodile". The specific name, colletti, honors Mr. Collett for the discovery of the holotype and only known specimen.
Eohyosaurus is an extinct genus of basal rhynchosaur known from the early Middle Triassic Burgersdorp Formation of Free State, South Africa. It contains a single species, Eohyosaurus wolvaardti.
Langeronyx is an extinct genus of basal rhynchosaurid known from the early Middle Triassic Bromsgrove Sandstone Formation of Warwickshire, UK. It contains a single species, Langeronyx brodiei, originally included in the genus Rhynchosaurus. R. brodiei was first described and named by Michael Benton in 1990, but its redescription by Martín D. Ezcurra, Felipe Montefeltro and Richard J. Butler in 2016 recovered it as more closely related to the more advance hyperodapedontine than to the type species of Rhynchosaurus and thus it was moved to its own genus. The generic name Langeronyx honors the Brazilian paleontologist Max Cardoso Langer in recognition of his rhynchosaur research, combined with the Greek onyx (óνυξ) meaning "claw", a common suffix for rhynchosaur genera. L. brodiei is known solely from the holotype, a partial skull divided into the two specimens WARMS G6097/1 and NHMUK PV R8495, housed in the Warwickshire Museum, Warwick and Natural History Museum, London, respectively. Other specimens originally referred to R. brodiei either do not overlap with its type or can be just as likely referred to other basal rhynchosaurids. L. brodiei is one of two basal archosauromorphs known from the Bromsgrove Sandstone Formation, the other being the lesser known Rhombopholis scutulata.

Azendohsauridae is a family of allokotosaurian archosauromorphs that lived during the Middle to Late Triassic period, around 242-216 million years ago. The family was originally named solely for the eponymous Azendohsaurus, marking out its distinctiveness from other allokotosaurs, but as of 2022 the family now includes four other genera: the basal genus Pamelaria, the large horned herbivore Shringasaurus, and two carnivorous genera grouped into the subfamily-level subclade Malerisaurinae, Malerisaurus and Puercosuchus, and potentially also the dubious genus Otischalkia. Most fossils of azendohsaurids have a Gondwanan distribution, with multiple species known across Morocco and Madagascar in Africa as well as India, although fossils of malerisaurine azendohsaurids have also been found in the southwestern United States of North America.
Oryctorhynchus is an extinct genus of rhynchosaur from the Late Triassic (Carnian-Norian)-aged Wolfville Formation of Nova Scotia, Canada that may have been the same animal as Beesiiwo. The type species, O. bairdi, was named and described in 2020. It was originally seen as a species of Hyperodapedon until 2020.







