Related Research Articles
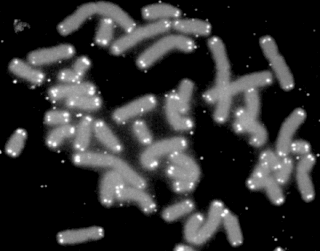
A telomere is a region of repetitive nucleotide sequences associated with specialized proteins at the ends of linear chromosomes. Although there are different architectures, telomeres, in a broad sense, are a widespread genetic feature most commonly found in eukaryotes. In most, if not all species possessing them, they protect the terminal regions of chromosomal DNA from progressive degradation and ensure the integrity of linear chromosomes by preventing DNA repair systems from mistaking the very ends of the DNA strand for a double strand break.

Telomerase, also called terminal transferase, is a ribonucleoprotein that adds a species-dependent telomere repeat sequence to the 3' end of telomeres. A telomere is a region of repetitive sequences at each end of the chromosomes of most eukaryotes. Telomeres protect the end of the chromosome from DNA damage or from fusion with neighbouring chromosomes. The fruit fly Drosophila melanogaster lacks telomerase, but instead uses retrotransposons to maintain telomeres.
Subtelomeres are segments of DNA between telomeric caps and chromatin.

In molecular biology, G-quadruplex secondary structures (G4) are formed in nucleic acids by sequences that are rich in guanine. They are helical in shape and contain guanine tetrads that can form from one, two or four strands. The unimolecular forms often occur naturally near the ends of the chromosomes, better known as the telomeric regions, and in transcriptional regulatory regions of multiple genes, both in microbes and across vertebrates including oncogenes in humans. Four guanine bases can associate through Hoogsteen hydrogen bonding to form a square planar structure called a guanine tetrad, and two or more guanine tetrads can stack on top of each other to form a G-quadruplex.
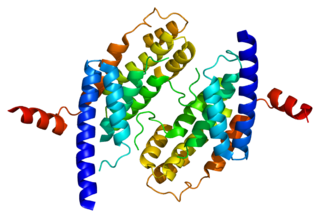
Telomeric repeat-binding factor 2 is a protein that is present at telomeres throughout the cell cycle. It is also known as TERF2, TRF2, and TRBF2, and is encoded in humans by the TERF2 gene. It is a component of the shelterin nucleoprotein complex and a second negative regulator of telomere length, playing a key role in the protective activity of telomeres. It was first reported in 1997 in the lab of Titia de Lange, where a DNA sequence similar, but not identical, to TERF1 was discovered, with respect to the Myb-domain. De Lange isolated the new Myb-containing protein sequence and called it TERF2.

Telomeric repeat-binding factor 1 is a protein that in humans is encoded by the TERF1 gene.

Telomerase RNA component, also known as TR, TER or TERC, is an ncRNA found in eukaryotes that is a component of telomerase, the enzyme used to extend telomeres. TERC serves as a template for telomere replication by telomerase. Telomerase RNAs differ greatly in sequence and structure between vertebrates, ciliates and yeasts, but they share a 5' pseudoknot structure close to the template sequence. The vertebrate telomerase RNAs have a 3' H/ACA snoRNA-like domain.

Protection of telomeres protein 1 is a protein that in humans is encoded by the POT1 gene.
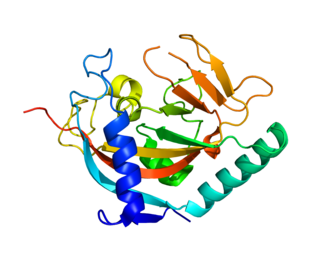
Tankyrase, also known as tankyrase 1, is an enzyme that in humans is encoded by the TNKS gene. It inhibits the binding of TERF1 to telomeric DNA. Tankyrase attracts substantial interest in cancer research through its interaction with AXIN1 and AXIN2, which are negative regulators of pro-oncogenic β-catenin signaling. Importantly, activity in the β-catenin destruction complex can be increased by tankyrase inhibitors and thus such inhibitors are a potential therapeutic option to reduce the growth of β-catenin-dependent cancers.

TERF1-interacting nuclear factor 2 is a protein that in humans is encoded by the TINF2 gene. TINF2 is a component of the shelterin protein complex found at the end of telomeres.

Adrenocortical dysplasia protein homolog is a protein that in humans is encoded by the ACD gene.

Telomeric repeat-binding factor 2-interacting protein 1 also known as repressor activator protein 1 (Rap1) is a protein that in humans is encoded by the TERF2IP gene.
PIN2/TERF1-interacting telomerase inhibitor 1, also known as PINX1, is a human gene. PINX1 is also known as PIN2 interacting protein 1. PINX1 is a telomerase inhibitor and a possible tumor suppressor.

Tankyrase-2 is an enzyme that in humans is encoded by the TNKS2 gene.
Alternative Lengthening of Telomeres is a telomerase-independent mechanism by which cancer cells avoid the degradation of telomeres.

A mega-telomere, is an extremely long telomere sequence that sits on the end of chromosomes and prevents the loss of genetic information during cell replication. Like regular telomeres, mega-telomeres are made of a repetitive sequence of DNA and associated proteins, and are located on the ends of chromosomes. However, mega-telomeres are substantially longer than regular telomeres, ranging in size from 50 kilobases to several megabases.
Shelterin is a protein complex known to protect telomeres in many eukaryotes from DNA repair mechanisms, as well as to regulate telomerase activity. In mammals and other vertebrates, telomeric DNA consists of repeating double-stranded 5'-TTAGGG-3' (G-strand) sequences along with the 3'-AATCCC-5' (C-strand) complement, ending with a 50-400 nucleotide 3' (G-strand) overhang. Much of the final double-stranded portion of the telomere forms a T-loop (Telomere-loop) that is invaded by the 3' (G-strand) overhang to form a small D-loop (Displacement-loop).

Titia de Lange is the Director of the Anderson Center for Cancer Research, the Leon Hess professor and the head of Laboratory Cell Biology and Genetics at Rockefeller University.

Telomeric repeat–containing RNA (TERRA) is a long non-coding RNA transcribed from telomeres - repetitive nucleotide regions found on the ends of chromosomes that function to protect DNA from deterioration or fusion with neighboring chromosomes. TERRA has been shown to be ubiquitously expressed in almost all cell types containing linear chromosomes - including humans, mice, and yeasts. While the exact function of TERRA is still an active area of research, it is generally believed to play a role in regulating telomerase activity as well as maintaining the heterochromatic state at the ends of chromosomes. TERRA interaction with other associated telomeric proteins has also been shown to help regulate telomere integrity in a length-dependent manner.
Telomeres, the caps on the ends of eukaryotic chromosomes, play critical roles in cellular aging and cancer. An important facet to how telomeres function in these roles is their involvement in cell cycle regulation.
References
- ↑ Database, GeneCards Human Gene. "TERF2 Gene - GeneCards | TERF2 Protein | TERF2 Antibody". www.genecards.org. Retrieved 2017-11-30.
- ↑ Lee WK, Cho MH (June 2016). "Telomere-binding protein regulates the chromosome ends through the interaction with histone deacetylases in Arabidopsis thaliana". Nucleic Acids Research. 44 (10): 4610–24. doi:10.1093/nar/gkw067. PMC 4889915 . PMID 26857545.
- ↑ Ilicheva NV, Podgornaya OI, Voronin AP (2015-01-01). Donev R (ed.). Advances in Protein Chemistry and Structural Biology. Vol. 101. Academic Press. pp. 67–96. doi:10.1016/bs.apcsb.2015.06.009. ISBN 9780128033678. PMID 26572976.
- 1 2 3 4 5 Sfeir A (September 2012). "Telomeres at a glance". Journal of Cell Science. 125 (Pt 18): 4173–8. doi:10.1242/jcs.106831. PMC 6518153 . PMID 23135002.
- 1 2 3 4 5 de Lange T (September 2005). "Shelterin: the protein complex that shapes and safeguards human telomeres". Genes & Development. 19 (18): 2100–10. doi: 10.1101/gad.1346005 . PMID 16166375.
- 1 2 3 Cookson JC, Laughton CA (March 2009). "The levels of telomere-binding proteins in human tumours and therapeutic implications". European Journal of Cancer. 45 (4): 536–50. doi:10.1016/j.ejca.2008.11.014. PMID 19114299.
- 1 2 3 Bianchi A, Smith S, Chong L, Elias P, de Lange T (April 1997). "TERF1 is a dimer and bends telomeric DNA". The EMBO Journal. 16 (7): 1785–94. doi:10.1093/emboj/16.7.1785. PMC 1169781 . PMID 9130722.
- 1 2 3 4 5 6 Smogorzewska A, van Steensel B, Bianchi A, Oelmann S, Schaefer MR, Schnapp G, de Lange T (March 2000). "Control of human telomere length by TERF1 and TERF2". Molecular and Cellular Biology. 20 (5): 1659–68. doi:10.1128/mcb.20.5.1659-1668.2000. PMC 85349 . PMID 10669743.
- ↑ He Q, Zeng P, Tan JH, Ou TM, Gu LQ, Huang ZS, Li D (July 2014). "G-quadruplex-mediated regulation of telomere binding protein POT1 gene expression". Biochimica et Biophysica Acta (BBA) - General Subjects. 1840 (7): 2222–33. doi:10.1016/j.bbagen.2014.03.001. PMID 24631651.
- 1 2 Sainger RN, Telang SD, Shukla SN, Patel PS (February 2007). "Clinical significance of telomere length and associated proteins in oral cancer". Biomarker Insights. 2: 9–19. doi:10.1177/117727190700200030. PMC 2717828 . PMID 19662189.
- ↑ van Steensel B, Smogorzewska A, de Lange T (February 1998). "TERF2 protects human telomeres from end-to-end fusions". Cell. 92 (3): 401–13. doi: 10.1016/s0092-8674(00)80932-0 . PMID 9476899.
- 1 2 3 4 5 Celli GB, de Lange T (July 2005). "DNA processing is not required for ATM-mediated telomere damage response after TERF2 deletion". Nature Cell Biology. 7 (7): 712–8. doi:10.1038/ncb1275. PMID 15968270.
- 1 2 3 Muñoz P, Blanco R, Flores JM, Blasco MA (October 2005). "XPF nuclease-dependent telomere loss and increased DNA damage in mice overexpressing TERF2 result in premature aging and cancer". Nature Genetics. 37 (10): 1063–71. doi:10.1038/ng1633. PMID 16142233.