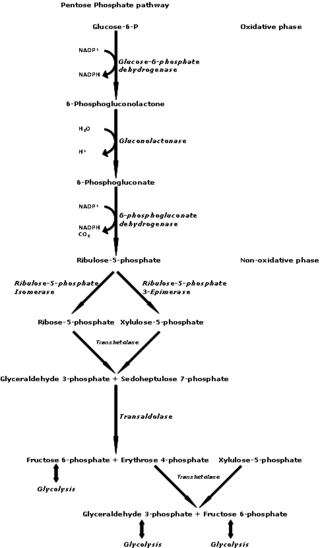
Transketolase (abbreviated as TK) is an enzyme that, in humans, is encoded by the TKTgene.[1] It participates in both the pentose phosphate pathway in all organisms and the Calvin cycle of photosynthesis. Transketolase catalyzes two important reactions, which operate in opposite directions in these two pathways. In the first reaction of the non-oxidative pentose phosphate pathway, the cofactor thiamine diphosphate accepts a 2-carbon fragment from a 5-carbon ketose (D-xylulose-5-P), then transfers this fragment to a 5-carbon aldose (D-ribose-5-P) to form a 7-carbon ketose (sedoheptulose-7-P). The abstraction of two carbons from D-xylulose-5-P yields the 3-carbon aldose glyceraldehyde-3-P. In the Calvin cycle, transketolase catalyzes the reverse reaction, the conversion of sedoheptulose-7-P and glyceraldehyde-3-P to pentoses, the aldose D-ribose-5-P and the ketose D-xylulose-5-P.
The second reaction catalyzed by transketolase in the pentose phosphate pathway involves the same thiamine diphosphate-mediated transfer of a 2-carbon fragment from D-xylulose-5-P to the aldose erythrose-4-phosphate, affording fructose 6-phosphate and glyceraldehyde-3-P. Again, in the Calvin cycle exactly the same reaction occurs, but in the opposite direction. Moreover, in the Calvin cycle this is the first reaction catalyzed by transketolase, rather than the second.
In mammals, transketolase connects the pentose phosphate pathway to glycolysis, feeding excess sugar phosphates into the main carbohydrate metabolic pathways. Its presence is necessary for the production of NADPH, especially in tissues actively engaged in biosyntheses, such as fatty acid synthesis by the liver and mammary glands, and for steroid synthesis by the liver and adrenal glands. Thiamine diphosphate is an essential cofactor, along with calcium.
Transketolase is abundantly expressed in the mammalian cornea by the stromal keratocytes and epithelial cells and is reputed to be one of the corneal crystallins.[2]
Species distribution
Transketolase is widely expressed in a wide range of organisms including bacteria, plants, and mammals. The following human genes encode proteins with transketolase activity:
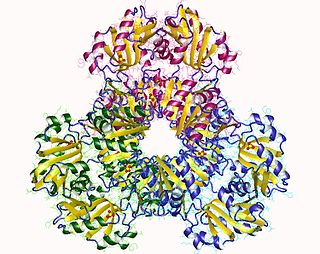
The entrance to the active site for this enzyme is made up mainly of several arginine, histidine, serine, and aspartate side-chains, with a glutamate side-chain playing a secondary role. These side-chains, to be specific Arg359, Arg528, His469, and Ser386, are conserved within each transketolase enzyme and interact with the phosphate group of the donor and acceptor substrates. Because the substrate channel is so narrow, the donor and acceptor substrates cannot be bound simultaneously. Also, the substrates conform into a slightly extended form upon binding in the active site to accommodate this narrow channel.
Although this enzyme is able to bind numerous types of substrates, such as phosphorylated and nonphosphorylated monosaccharides including the keto and aldosugars fructose, ribose, etc., it has a high specificity for the stereoconfiguration of the hydroxyl groups of the sugars. These hydroxyl groups at C-3 and C-4 of the ketose donor must be in the D-threo configuration in order to correctly correspond to the C-1 and C-2 positions on the aldose acceptor.[3] Also they stabilize the substrate in the active site by interacting with the Asp477, His30, and His263 residues. Disruption of this configuration, both the placement of hydroxyl groups or their stereochemistry, would consequently alter the H-bonding between the residues and substrates thus causing a lower affinity for the substrates.
In the first half of this pathway, His263 is used to effectively abstract the C3 hydroxyl proton, which thus allows a 2-carbon segment to be cleaved from fructose 6-phosphate.[4] The cofactor necessary for this step to occur is thiamin pyrophosphate (TPP). The binding of TPP to the enzyme incurs no major conformational change to the enzyme; instead, the enzyme has two flexible loops at the active site that make TPP accessible and binding possible.[3] Thus, this allows the active site to have a "closed" conformation rather than a large conformational change. Later in the pathway, His263 is used as a proton donor for the substrate acceptor-TPP complex, which can then generate erythrose-4-phosphate.
The histidine and aspartate side-chains are used to effectively stabilize the substrate within the active site and also participate in deprotonation of the substrate. To be specific, the His 263 and His30 side-chains form hydrogen bonds to the aldehyde end of the substrate, which is deepest into the substrate channel, and Asp477 forms hydrogen bonds with the alpha hydroxyl group on the substrate, where it works to effectively bind the substrate and check for proper stereochemistry. It is also thought that Asp477 could have important catalytic effects because of its orientation in the middle of the active site and its interactions with the alpha hydroxyl group of the substrate. Glu418, which is located in the deepest region of the active site, plays a critical role in stabilizing the TPP cofactor. To be specific, it is involved in the cofactor-assisted proton abstraction from the substrate molecule.[3]
The phosphate group of the substrate also plays an important role in stabilizing the substrate upon its entrance into the active site. The tight ionic and polar interactions between this phosphate group and the residues Arg359, Arg528, His469, and Ser386 collectively work to stabilize the substrate by forming H-bonds to the oxygen atoms of the phosphate.[3] The ionic nature is found in the salt bridge formed from Arg359 to the phosphate group.
Mechanism
The catalysis of this mechanism is initiated by the deprotonation of TPP at the thiazolium ring. This carbanion then binds to the carbonyl of the donor substrate thus cleaving the bond between C-2 and C-3. This keto fragment remains covalently bound to the C-2 carbon of TPP. The donor substrate is then released, and the acceptor substrate enters the active site where the fragment, which is bound to the intermediate α-β-dihydroxyethyl thiamin diphosphate, is then transferred to the acceptor.[3]
Experiments have also been conducted that test the effect replacing alanine for the amino acids at the entrance to the active site, Arg359, Arg528, and His469, which interact with the phosphate group of the substrate. This replacement creates a mutant enzyme with impaired catalytic activity.[3]
In Wernicke–Korsakoff syndrome, while no mutations could be demonstrated,[6] there is an indication that thiamine deficiency leads to Wernicke–Korsakoff syndrome only in those whose transketolase has a reduced affinity for thiamine.[7] In this way, the activity of transketolase is greatly hindered, and, as a consequence, the entire pentose phosphate pathway is inhibited.[8]
In Transketolase Deficiency, also known as SDDHD (Short Stature, Developmental Delay, and congenital Heart Defects), the disease is caused by an inherited autosomal recessive mutation in the TKT gene. A rare disorder of pentose phosphate metabolism with symptoms apparent in infancy including developmental delay and intellectual disability, delayed or absent speech, short stature, and congenital heart defects. Additional reported features include hypotonia, hyperactivity, stereotypic behavior, ophthalmologic abnormalities, hearing impairment, and variable facial dysmorphism, among others. Laboratory analysis shows elevated plasma and urinary polyols (erythritol, arabitol, and ribitol) and urinary sugar-phosphates (ribose-5-phosphate and xylulose/ribulose-5-phosphate).[9] "Cell extracts from all 5 patients showed absent or low residual TKT activity. Boyle et al. (2016) suggested that the low TKT activity in some tissues, possibly from another protein with the same function, might explain why TKT deficiency is compatible with life even though TKT is an essential enzyme."[10]
Diagnostic use
Red blood cell transketolase activity is reduced in deficiency of thiamine (vitamin B1), and may be used in the diagnosis of Wernicke encephalopathy and other B1-deficiency syndromes if the diagnosis is in doubt.[11] Apart from the baseline enzyme activity (which may be normal even in deficiency states), acceleration of enzyme activity after the addition of thiamine pyrophosphate may be diagnostic of thiamine deficiency (0-15% normal, 15-25% deficiency, >25% severe deficiency).[12]
Related Research Articles
Thiamine, also known as thiamin and vitamin B1, is a vitamin, an essential micronutrient for humans and animals. It is found in food and commercially synthesized to be a dietary supplement or medication. Phosphorylated forms of thiamine are required for some metabolic reactions, including the breakdown of glucose and amino acids.
Isomerases are a general class of enzymes that convert a molecule from one isomer to another. Isomerases facilitate intramolecular rearrangements in which bonds are broken and formed. The general form of such a reaction is as follows:
A tetrose is a monosaccharide with 4 carbon atoms. They have either an aldehyde functional group in position 1 (aldotetroses) or a ketone functional group in position 2 (ketotetroses).
Thiamine pyrophosphate (TPP or ThPP), or thiamine diphosphate (ThDP), or cocarboxylase is a thiamine (vitamin B1) derivative which is produced by the enzyme thiamine diphosphokinase. Thiamine pyrophosphate is a cofactor that is present in all living systems, in which it catalyzes several biochemical reactions.
The pentose phosphate pathway is a metabolic pathway parallel to glycolysis. It generates NADPH and pentoses as well as ribose 5-phosphate, a precursor for the synthesis of nucleotides. While the pentose phosphate pathway does involve oxidation of glucose, its primary role is anabolic rather than catabolic. The pathway is especially important in red blood cells (erythrocytes).
Ribulose is a ketopentose — a monosaccharide containing five carbon atoms, and including a ketone functional group. It has chemical formula C5H10O5. Two enantiomers are possible, d-ribulose and l-ribulose. d-Ribulose is the diastereomer of d-xylulose.
Pyruvate decarboxylase is an enzyme that catalyses the decarboxylation of pyruvic acid to acetaldehyde. It is also called 2-oxo-acid carboxylase, alpha-ketoacid carboxylase, and pyruvic decarboxylase. In anaerobic conditions, this enzyme is participates in the fermentation process that occurs in yeast, especially of the genus Saccharomyces, to produce ethanol by fermentation. It is also present in some species of fish where it permits the fish to perform ethanol fermentation when oxygen is scarce. Pyruvate decarboxylase starts this process by converting pyruvate into acetaldehyde and carbon dioxide. Pyruvate decarboxylase depends on cofactors thiamine pyrophosphate (TPP) and magnesium. This enzyme should not be mistaken for the unrelated enzyme pyruvate dehydrogenase, an oxidoreductase, that catalyzes the oxidative decarboxylation of pyruvate to acetyl-CoA.
Sugar phosphates are often used in biological systems to store or transfer energy. They also form the backbone for DNA and RNA. Sugar phosphate backbone geometry is altered in the vicinity of the modified nucleotides.
In enzymology, aldose reductase is a cytosolic NADPH-dependent oxidoreductase that catalyzes the reduction of a variety of aldehydes and carbonyls, including monosaccharides. It is primarily known for catalyzing the reduction of glucose to sorbitol, the first step in polyol pathway of glucose metabolism.
Transaldolase is an enzyme of the non-oxidative phase of the pentose phosphate pathway. In humans, transaldolase is encoded by the TALDO1 gene.
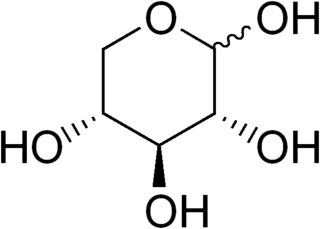
Ribose 5-phosphate (R5P) is both a product and an intermediate of the pentose phosphate pathway. The last step of the oxidative reactions in the pentose phosphate pathway is the production of ribulose 5-phosphate. Depending on the body's state, ribulose 5-phosphate can reversibly isomerize to ribose 5-phosphate. Ribulose 5-phosphate can alternatively undergo a series of isomerizations as well as transaldolations and transketolations that result in the production of other pentose phosphates as well as fructose 6-phosphate and glyceraldehyde 3-phosphate.
D-Xylulose 5-phosphate (D-xylulose-5-P) is an intermediate in the pentose phosphate pathway. It is a ketose sugar formed from ribulose-5-phosphate by ribulose-5-phosphate epimerase. In the non-oxidative branch of the pentose phosphate pathway, xylulose-5-phosphate acts as a donor of two-carbon ketone groups in transketolase reactions.
Phosphopentose epimerase encoded by the RPE gene is a metalloprotein that catalyzes the interconversion between D-ribulose 5-phosphate and D-xylulose 5-phosphate.
Ribose-phosphate diphosphokinase is an enzyme that converts ribose 5-phosphate into phosphoribosyl pyrophosphate (PRPP). It is classified under EC 2.7.6.1.
D-Xylose is a five-carbon aldose that can be catabolized or metabolized into useful products by a variety of organisms.
Ribose-5-phosphate isomerase (Rpi) encoded by the RPIA gene is an enzyme that catalyzes the conversion between ribose-5-phosphate (R5P) and ribulose-5-phosphate (Ru5P). It is a member of a larger class of isomerases which catalyze the interconversion of chemical isomers. It plays a vital role in biochemical metabolism in both the pentose phosphate pathway and the Calvin cycle. The systematic name of this enzyme class is D-ribose-5-phosphate aldose-ketose-isomerase.
In enzymology, a S-methyl-5-thioribose-1-phosphate isomerase is an enzyme that catalyzes the chemical reaction
Diphosphomevalonate decarboxylase (EC 4.1.1.33), most commonly referred to in scientific literature as mevalonate diphosphate decarboxylase, is an enzyme that catalyzes the chemical reaction
The enzyme phosphoketolase(EC 4.1.2.9) catalyzes the chemical reactions
In enzymology, a formaldehyde transketolase is an enzyme that catalyzes the chemical reaction
↑ Wikner C, Nilsson U, Meshalkina L, Udekwu C, Lindqvist Y, Schneider G (December 1997). "Identification of catalytically important residues in yeast transketolase". Biochemistry. 36 (50): 15643–9. doi:10.1021/bi971606b. PMID9398292.
↑ Tabarki B, Al-Hashem A, Alfadhel M (August 2020). "Biotin-Thiamine-Responsive Basal Ganglia Disease". In Adam MP, Ardinger HH, Pagon RA, Wallace SE, Bean LJ, Mirzaa G, Amemiya A, Tabarki B, Al-Hashem A, Alfadhel M (eds.). GeneReviews. University of Washington, Seattle. PMID24260777.
↑ Blass JP, Gibson GE (December 1977). "Abnormality of a thiamine-requiring enzyme in patients with Wernicke-Korsakoff syndrome". The New England Journal of Medicine. 297 (25): 1367–70. doi:10.1056/NEJM197712222972503. PMID927453.
↑ Doolman R, Dinbar A, Sela BA (July 1995). "Improved measurement of transketolase activity in the assessment of "TPP effect"". European Journal of Clinical Chemistry and Clinical Biochemistry. 33 (7): 445–6. PMID7548453.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.