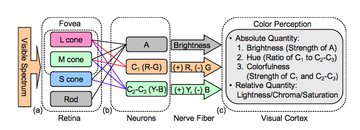
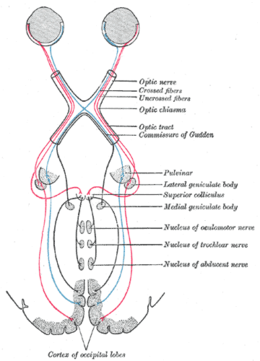
Color or colour is the visual perception based on the electromagnetic spectrum. Though color is not an inherent property of matter, color perception is related to an object's light absorption, reflection, emission spectra and interference. For most humans, colors are perceived in the visible light spectrum with three types of cone cells (trichromacy). Other animals may have a different number of cone cell types or have eyes sensitive to different wavelength, such as bees that can distinguish ultraviolet, and thus have a different color sensitivity range. Animal perception of color originates from different light wavelength or spectral sensitivity in cone cell types, which is then processed by the brain.

Color blindness or color vision deficiency (CVD) is the decreased ability to see color or differences in color. The severity of color blindness ranges from mostly unnoticeable to full absence of color perception. Color blindness is usually an inherited problem or variation in the functionality of one or more of the three classes of cone cells in the retina, which mediate color vision. The most common form is caused by a genetic condition called congenital red–green color blindness, which affects up to 1 in 12 males (8%) and 1 in 200 females (0.5%). The condition is more prevalent in males, because the opsin genes responsible are located on the X chromosome. Rarer genetic conditions causing color blindness include congenital blue–yellow color blindness, blue cone monochromacy, and achromatopsia. Color blindness can also result from physical or chemical damage to the eye, the optic nerve, parts of the brain, or from medication toxicity. Color vision also naturally degrades in old age.

A set of primary colors or primary colours consists of colorants or colored lights that can be mixed in varying amounts to produce a gamut of colors. This is the essential method used to create the perception of a broad range of colors in, e.g., electronic displays, color printing, and paintings. Perceptions associated with a given combination of primary colors can be predicted by an appropriate mixing model that reflects the physics of how light interacts with physical media, and ultimately the retina.

In color theory, hue is one of the main properties of a color, defined technically in the CIECAM02 model as "the degree to which a stimulus can be described as similar to or different from stimuli that are described as red, orange, yellow, green, blue, violet," within certain theories of color vision.

The Natural Color System (NCS) is a proprietary perceptual color model. It is based on the color opponency hypothesis of color vision, first proposed by German physiologist Ewald Hering. The current version of the NCS was developed by the Swedish Colour Centre Foundation, from 1964 onwards. The research team consisted of Anders Hård, Lars Sivik and Gunnar Tonnquist, who in 1997 received the AIC Judd award for their work. The system is based entirely on the phenomenology of human perception and not on color mixing. It is illustrated by a color atlas, marketed by NCS Colour AB in Stockholm.

Color constancy is an example of subjective constancy and a feature of the human color perception system which ensures that the perceived color of objects remains relatively constant under varying illumination conditions. A green apple for instance looks green to us at midday, when the main illumination is white sunlight, and also at sunset, when the main illumination is red. This helps us identify objects.

Color vision, a feature of visual perception, is an ability to perceive differences between light composed of different frequencies independently of light intensity.

Tetrachromacy is the condition of possessing four independent channels for conveying color information, or possessing four types of cone cell in the eye. Organisms with tetrachromacy are called tetrachromats.

An afterimage is an image that continues to appear in the eyes after a period of exposure to the original image. An afterimage may be a normal phenomenon or may be pathological (palinopsia). Illusory palinopsia may be a pathological exaggeration of physiological afterimages. Afterimages occur because photochemical activity in the retina continues even when the eyes are no longer experiencing the original stimulus.

Trichromacy or trichromatism is the possession of three independent channels for conveying color information, derived from the three different types of cone cells in the eye. Organisms with trichromacy are called trichromats.
Dichromacy is the state of having two types of functioning photoreceptors, called cone cells, in the eyes. Organisms with dichromacy are called dichromats. Dichromats require only two primary colors to be able to represent their visible gamut. By comparison, trichromats need three primary colors, and tetrachromats need four. Likewise, every color in a dichromat's gamut can be evoked with monochromatic light. By comparison, every color in a trichromat's gamut can be evoked with a combination of monochromatic light and white light.
The opponent process is a color theory that states that the human visual system interprets information about color by processing signals from photoreceptor cells in an antagonistic manner. The opponent-process theory suggests that there are three opponent channels, each comprising an opposing color pair: red versus green, blue versus yellow, and black versus white (luminance). The theory was first proposed in 1892 by the German physiologist Ewald Hering.
A color model is an abstract mathematical model describing the way colors can be represented as tuples of numbers, typically as three or four values or color components. When this model is associated with a precise description of how the components are to be interpreted, taking account of visual perception, the resulting set of colors is called "color space."

The Abney effect or the purity-on-hue effect describes the perceived hue shift that occurs when white light is added to a monochromatic light source.

Infant vision concerns the development of visual ability in human infants from birth through the first years of life. The aspects of human vision which develop following birth include visual acuity, tracking, color perception, depth perception, and object recognition.

Chromostereopsis is a visual illusion whereby the impression of depth is conveyed in two-dimensional color images, usually of red–blue or red–green colors, but can also be perceived with red–grey or blue–grey images. Such illusions have been reported for over a century and have generally been attributed to some form of chromatic aberration.

Impossible colors are colors that do not appear in ordinary visual functioning. Different color theories suggest different hypothetical colors that humans are incapable of perceiving for one reason or another, and fictional colors are routinely created in popular culture. While some such colors have no basis in reality, phenomena such as cone cell fatigue enable colors to be perceived in certain circumstances that would not be otherwise.

A parasol cell, sometimes called an M cell or M ganglion cell, is one type of retinal ganglion cell (RGC) located in the ganglion cell layer of the retina. These cells project to magnocellular cells in the lateral geniculate nucleus (LGN) as part of the magnocellular pathway in the visual system. They have large cell bodies as well as extensive branching dendrite networks and as such have large receptive fields. Relative to other RGCs, they have fast conduction velocities. While they do show clear center-surround antagonism, they receive no information about color. Parasol ganglion cells contribute information about the motion and depth of objects to the visual system.

Russell L. De Valois was an American scientist recognized for his pioneering research on spatial and color vision.

The memory color effect is the phenomenon that the canonical hue of a type of object acquired through experience can directly modulate the appearance of the actual colors of objects.