Embryomics is the identification, characterization and study of the diverse cell types which arise during embryogenesis, especially as this relates to the location and developmental history of cells in the embryo. Cell type may be determined according to several criteria: location in the developing embryo, gene expression as indicated by protein and nucleic acid markers and surface antigens, and also position on the embryogenic tree.
There are many cell markers useful in distinguishing, classifying, separating and purifying the numerous cell types present at any given time in a developing organism. These cell markers consist of select RNAs and proteins present inside, and surface antigens present on the surface of, the cells making up the embryo. For any given cell type, these RNA and protein markers reflect the genes characteristically active in that cell type. The catalog of all these cell types and their characteristic markers is known as the organism's embryome.[citation needed] The word is a portmanteau of embryo and genome. “Embryome” may also refer to the totality of the physical cell markers themselves.
As an embryo develops from a fertilized egg, the single egg cell splits into many cells, which grow in number and migrate to the appropriate locations inside the embryo at appropriate times during development. As the embryo's cells grow in number and migrate, they also differentiate into an increasing number of different cell types, ultimately turning into the stable, specialized cell types characteristic of the adult organism. Each of the cells in an embryo contains the same genome, characteristic of the species,[1] but the level of activity of each of the many thousands of genes that make up the complete genome varies with, and determines, a particular cell's type (e.g. neuron, bone cell, skin cell, muscle cell, etc.).
During embryo development (embryogenesis), many cell types are present which are not present in the adult organism. These temporary cells are called progenitor cells, and are intermediate cell types which disappear during embryogenesis by turning into other progenitor cells, or into mature adult somatic cell types, or which disappear due to programmed cell death (apoptosis).
The entire process of embryogenesis can be described with the aid of two maps: an embryo map, a temporal sequence of 3-dimensional images of the developing embryo, showing the location of cells of the many cell types present in the embryo at a given time, and an embryogenic tree, a diagram showing how the cell types are derived from each other during embryogenesis.
The embryo map is a sequence of 3-D images, or slices of 3-D images, of the developing embryo which, if viewed rapidly in temporal order, forms a time-lapse view of the growing embryo.
The embryogenic tree is a diagram which shows the temporal development of each of the cell lines in the embryo. When drawn on a piece of paper, this diagram takes the form of a tree, analogous to the evolutionary tree of life which illustrates the development of life on Earth. However, instead of each branch on this tree representing a species, as in the tree of life, each branch represents a particular cell type present in the embryo at a particular time. And of course, an embryogenic tree covers the gestation period of weeks or months, instead of billions of years, as in the case of the evolutionary tree of life.
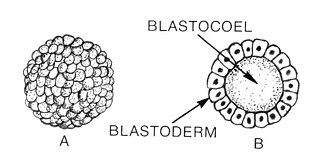
Human embryogenesis is the referent here, but embryogenesis in other vertebrate species closely follows the same pattern. The egg cell (ovum), after fertilization with a sperm cell, becomes the zygote, represented by the trunk at the very bottom of the tree. This single zygote cell divides in two, three times, forming first a cluster of two-cells, then four-cells, and finally eight-cells. One more cell division brings the number of cells to 16, at which time it is called a morula, instead of a zygote. This ball of 16 cells then reorganizes into a hollow sphere called a blastocyst. As the number of cells grows from 16 to between 40 and 150, the blastocyst differentiates into two layers, an outer sphere of cells called the trophoblast and an inner cell mass called the embryoblast.
The spherical outer cell layer (trophoblast), after implantation in the wall of the uterus, further differentiates and grows to form the placenta.
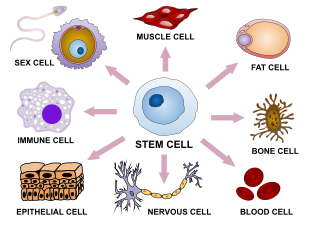
The cells of the inner cell mass (embryoblast), which are known as human embryonic stem cells (hESCs), will further differentiate to form four structures: the amnion, the yolk sac, the allantois, and the embryo itself. Human embryonic stem cells are pluripotent, that is, they can differentiate into any of the cell types present in the adult human, and into any of the intermediate progenitor cell types that eventually turn into the adult cell lines. hESCs are also immortal, in that they can divide and grow in number indefinitely, without undergoing either differentiation or cellular aging (cellular senescence).
The first differentiation of the hESCs that form the embryo proper, is into three cell types known as the germ layers: the ectoderm, the mesoderm, and the endoderm. The ectoderm eventually forms the skin (including hair and nails), mucous membranes and nervous system. The mesoderm forms the skeleton and muscles, heart and circulatory system, urinary and reproductive systems, and connective tissues inside the body. The endoderm forms the gastrointestinal tract (stomach and intestines), the respiratory tract, and the endocrine system (liver and endocrine glands).
Mapping the embryogenic tree
A primary goal in embryomics is a complete mapping the embryogenic tree: Identifying each of the cell types present in the developing embryo and placing it in the tree on its proper branch. There is an unknown number, probably thousands, of distinct cell types present in the developing embryo, including progenitor cell lines which are only temporarily present and which disappear either by differentiating into the permanent somatic cell types which make up the tissues of the infant's body at birth (or into other progenitor cell lines), or by undergoing the programmed cell death process known as apoptosis.
Each cell type is defined by which genes are characteristically active in that cell type. A particular gene in a cell's genome codes for the production of a particular protein, that is, when that gene is turned on (active), the protein coded for by that gene is produced and present somewhere in the cell. Production of a particular protein involves the production of a particular mRNA (messenger RNA) sequence as an intermediate step in protein synthesis. This mRNA is produced by copying process called transcription, from the DNA in the cell's nucleus. The mRNA so produced travels from the nucleus into the cytoplasm, where it encounters and latches onto ribosomes stuck to the cytoplasmic side of the endoplasmic reticulum. Attachment of the mRNA strand to the ribosome initiates the production of the protein coded for by the mRNA strand. Therefore, the profile of active genes in a cell is reflected in the presence or absence of corresponding proteins and mRNA strands in the cell's cytoplasm, and antigen proteins present on the cell's outer membrane. Discovering, determining and classifying cells as to their type therefore involves detecting and measuring the type and amount of specific protein and RNA molecules present in the cells.
In addition, mapping the tree of embryogenesis involves assigning to each specific, identifiable cell type, a particular branch, or place, in the tree. This requires knowing the “ancestry” of each cell type, that is, which cell type preceded it in the development process. This information can be deduced by observing in detail the distribution and placement of cells, by type, in the developing embryo, and by also observing, in cells growing in culture (“in vitro”) any differentiation events, should they occur for whatever reason, and by other means.
Cells, embryonic cells in particular, are sensitive to the presence or absence of specific chemical molecules in their surroundings. This is the basis for cell signaling, and during embryogenesis cells “talk to each other” by emitting and receiving signalling molecules. This is how development of the embryo's structure is organized and controlled. If cells of a particular line have been removed from the embryo and are growing alone in a Petri dish in the lab, and some cell signaling chemicals are put in the growth medium bathing the cells, this can induce the cells to differentiate into a different, “daughter” cell type, mimicking the differentiation process that occurs naturally in the developing embryo. Artificially inducing differentiation in this way can yield clues to the correct placement of a particular cell line in the embryogenic tree, by observing what kind of cell results from inducing the differentiation.
In the laboratory, human embryonic stem cells growing in culture can be induced to differentiate into progenitor cells by exposing the hESCs to chemicals (e.g. protein growth and differentiation factors) present in the developing embryo. The progenitor cells so produced may then be isolated into pure colonies, grown in culture, and then classified according to type and assigned positions in the embryogenic tree. Such purified cultures of progenitor cells may be used in research to study disease processes in vitro, as diagnostic tools, or potentially developed for use in regenerative medicine therapies.[2]
Regenerative medicine
Embryomics is the core science supporting the development of regenerative medicine. Regenerative medicine involves use of specially grown cells, tissues and organs as therapeutic agents to cure disease and repair injury, and springs from the development of mammalian cloning technology.[3] Other medical and surgical methods may use chemicals (pharmaceuticals) as therapeutic agents, or involve removal of injured or diseased tissue (surgery), or use inserted tissues or organs (transplant surgery). Use of transplanted tissue or organs in medicine is not classified as regenerative medicine, because the tissues and organs were not grown specifically for use as therapeutic agents.
Ultimately, one of the goals of regenerative medicine and applied embryomics, is the creation of cells, tissues and organs grown from cells taken from the patient to be treated. This would be accomplished by reprogramming adult stem or somatic cells removed from the patient, so that these cells revert to the pluripotent, embryonic state.[4][5][6] These synthetic stem cells would then be grown in culture and differentiated into the appropriate cell type indicated for treating the patient's disease or injury. The advantages here over current therapies are: elimination of immune rejection accompanying allograft transplantation, creation of a full complement of cells, tissues and organs as needed, and creation of youthful cells, tissues and organs for transplant and rejuvenation.
Technology for growing cells, tissues and organs for use in regenerative medicine can be developed by using the natural course of development of those cells, tissues and organs during embryogenesis, as a guide. Therefore, detailed knowledge of the complete embryome and the embryogenic tree is key to developing the full potential of regenerative medicine.
Embryomics also includes the application of embryomic data and theory, to the development of practical methods for evaluating, classifying, culturing, purifying, differentiating and manipulating human embryonic cells.
Notes
↑ The genome of course varies slightly from individual to individual, but these small variations are subsumed within the species genome.
↑ West, Michael D; Sargent, R Geoffrey; Long, Jeff; Brown, Colleen; Chu, Jing Song; Kessler, Steven; Derugin, Nikita; Sampathkumar, Janani; Burrows, Courtney; Vaziri, Homayoun; Williams, Roy; Chapman, Karen B; Larocca, David; Loring, Jeanne F; Murai, James (2008). "The ACTCellerate initiative: Large-scale combinatorial cloning of novel human embryonic stem cell derivatives". Regenerative Medicine. 3 (3): 287–308. doi:10.2217/17460751.3.3.287. PMID18462054.
↑ Wilmut, I.; Schnieke, A. E.; McWhir, J.; Kind, A. J.; Campbell, K. H. S. (1997). "Viable offspring derived from fetal and adult mammalian cells". Nature. 385 (6619): 810–3. doi:10.1038/385810a0. PMID9039911.
↑ Takahashi, Kazutoshi; Tanabe, Koji; Ohnuki, Mari; Narita, Megumi; Ichisaka, Tomoko; Tomoda, Kiichiro; Yamanaka, Shinya (2007). "Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors". Cell. 131 (5): 861–72. doi:10.1016/j.cell.2007.11.019. hdl:2433/49782. PMID18035408.
↑ Yu, J.; Vodyanik, M. A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J. L.; Tian, S.; Nie, J.; Jonsdottir, G. A.; Ruotti, V.; Stewart, R.; Slukvin, I. I.; Thomson, J. A. (2007). "Induced Pluripotent Stem Cell Lines Derived from Human Somatic Cells". Science. 318 (5858): 1917–20. doi:10.1126/science.1151526. PMID18029452.
Related Research Articles
Developmental biology is the study of the process by which animals and plants grow and develop. Developmental biology also encompasses the biology of regeneration, asexual reproduction, metamorphosis, and the growth and differentiation of stem cells in the adult organism.
In multicellular organisms, stem cells are undifferentiated or partially differentiated cells that can differentiate into various types of cells and divide indefinitely to produce more of the same stem cell. They are the earliest type of cell in a cell lineage. They are found in both embryonic and adult organisms, but they have slightly different properties in each. They are usually distinguished from progenitor cells, which cannot divide indefinitely, and precursor or blast cells, which are usually committed to differentiating into one cell type.
Cellular differentiation is the process in which a cell changes from one cell type to another. Usually, the cell changes to a more specialized type. Differentiation occurs numerous times during the development of a multicellular organism as it changes from a simple zygote to a complex system of tissues and cell types. Differentiation continues in adulthood as adult stem cells divide and create fully differentiated daughter cells during tissue repair and during normal cell turnover. Some differentiation occurs in response to antigen exposure. Differentiation dramatically changes a cell's size, shape, membrane potential, metabolic activity, and responsiveness to signals. These changes are largely due to highly controlled modifications in gene expression and are the study of epigenetics. With a few exceptions, cellular differentiation almost never involves a change in the DNA sequence itself. Thus, different cells can have very different physical characteristics despite having the same genome.
The blastula is a hollow sphere of cells, referred to as blastomeres, surrounding an inner fluid-filled cavity called the blastocoele formed during an early stage of embryonic development in animals. Embryo development begins with a sperm fertilizing an egg to become a zygote which undergoes many cleavages to develop into a ball of cells called a morula. Only when the blastocoele is formed does the early embryo become a blastula. The blastula precedes the formation of the gastrula in which the germ layers of the embryo form.
Plant callus is a growing mass of unorganized plant parenchyma cells. In living plants, callus cells are those cells that cover a plant wound. In biological research and biotechnology callus formation is induced from plant tissue samples (explants) after surface sterilization and plating onto tissue culture medium in vitro. The culture medium is supplemented with plant growth regulators, such as auxins, cytokinins, and gibberellins, to initiate callus formation or somatic embryogenesis. Callus initiation has been described for all major groups of land plants.
Embryonic stem cells are pluripotent stem cells derived from the inner cell mass of a blastocyst, an early-stage pre-implantation embryo. Human embryos reach the blastocyst stage 4–5 days post fertilization, at which time they consist of 50–150 cells. Isolating the embryoblast, or inner cell mass (ICM) results in destruction of the blastocyst, a process which raises ethical issues, including whether or not embryos at the pre-implantation stage should have the same moral considerations as embryos in the post-implantation stage of development.
Oct-4, also known as POU5F1, is a protein that in humans is encoded by the POU5F1 gene. Oct-4 is a homeodomain transcription factor of the POU family. It is critically involved in the self-renewal of undifferentiated embryonic stem cells. As such, it is frequently used as a marker for undifferentiated cells. Oct-4 expression must be closely regulated; too much or too little will cause differentiation of the cells.
Regenerative medicine is a branch of translational research in tissue engineering and molecular biology which deals with the "process of replacing, engineering or regenerating human or animal cells, tissues or organs to restore or establish normal function". This field holds the promise of engineering damaged tissues and organs by stimulating the body's own repair mechanisms to functionally heal previously irreparable tissues or organs.
Adult stem cells are undifferentiated cells, found throughout the body after development, that multiply by cell division to replenish dying cells and regenerate damaged tissues. Also known as somatic stem cells , they can be found in juvenile as well as adult animals and humans, unlike embryonic stem cells.
A stem cell line is a group of stem cells that is cultured in vitro and can be propagated indefinitely. Stem cell lines are derived from either animal or human tissues and come from one of three sources: embryonic stem cells, adult stem cells, or induced stem cells. They are commonly used in research and regenerative medicine.
In biology, reprogramming refers to erasure and remodeling of epigenetic marks, such as DNA methylation, during mammalian development or in cell culture. Such control is also often associated with alternative covalent modifications of histones.
The stem cell controversy is the consideration of the ethics of research involving the development and use of human embryos. Most commonly, this controversy focuses on embryonic stem cells. Not all stem cell research involves human embryos. For example, adult stem cells, amniotic stem cells, and induced pluripotent stem cells do not involve creating, using, or destroying human embryos, and thus are minimally, if at all, controversial. Many less controversial sources of acquiring stem cells include using cells from the umbilical cord, breast milk, and bone marrow, which are not pluripotent.
Endothelial stem cells (ESCs) are one of three types of stem cells found in bone marrow. They are multipotent, which describes the ability to give rise to many cell types, whereas a pluripotent stem cell can give rise to all types. ESCs have the characteristic properties of a stem cell: self-renewal and differentiation. These parent stem cells, ESCs, give rise to progenitor cells, which are intermediate stem cells that lose potency. Progenitor stem cells are committed to differentiating along a particular cell developmental pathway. ESCs will eventually produce endothelial cells (ECs), which create the thin-walled endothelium that lines the inner surface of blood vessels and lymphatic vessels.
Induced pluripotent stem cells are a type of pluripotent stem cell that can be generated directly from a somatic cell. The iPSC technology was pioneered by Shinya Yamanaka’s lab in Kyoto, Japan, who showed in 2006 that the introduction of four specific genes encoding transcription factors could convert somatic cells into pluripotent stem cells. He was awarded the 2012 Nobel Prize along with Sir John Gurdon "for the discovery that mature cells can be reprogrammed to become pluripotent."
Lin-28 homolog A is a protein that in humans is encoded by the LIN28 gene.
Cell potency is a cell's ability to differentiate into other cell types. The more cell types a cell can differentiate into, the greater its potency. Potency is also described as the gene activation potential within a cell, which like a continuum, begins with totipotency to designate a cell with the most differentiation potential, pluripotency, multipotency, oligopotency, and finally unipotency.
Somatic embryogenesis is an artificial process in which a plant or embryo is derived from a single somatic cell. Somatic embryos are formed from plant cells that are not normally involved in the development of embryos, i.e. ordinary plant tissue. No endosperm or seed coat is formed around a somatic embryo.
Induced stem cells (iSC) are stem cells derived from somatic, reproductive, pluripotent or other cell types by deliberate epigenetic reprogramming. They are classified as either totipotent (iTC), pluripotent (iPSC) or progenitor or unipotent – (iUSC) according to their developmental potential and degree of dedifferentiation. Progenitors are obtained by so-called direct reprogramming or directed differentiation and are also called induced somatic stem cells.
Directed differentiation is a bioengineering methodology at the interface of stem cell biology, developmental biology and tissue engineering. It is essentially harnessing the potential of stem cells by constraining their differentiation in vitro toward a specific cell type or tissue of interest. Stem cells are by definition pluripotent, able to differentiate into several cell types such as neurons, cardiomyocytes, hepatocytes, etc. Efficient directed differentiation requires a detailed understanding of the lineage and cell fate decision, often provided by developmental biology.
Musashi-2, also known as Musashi RNA binding protein 2, is a protein that in humans is encoded by the MSI2 gene. Like its homologue musashi-1 (MSI1), it is an RNA-binding protein involved in stemness.
References
West, Michael D. (September 16, 2003). The Immortal Cell (1st ed.). Doubleday.
Pansky, Ben (December 1, 1982). Review of Medical Embryology. Macmillan.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.