
In ecology, a niche is the match of a species to a specific environmental condition. It describes how an organism or population responds to the distribution of resources and competitors and how it in turn alters those same factors. "The type and number of variables comprising the dimensions of an environmental niche vary from one species to another [and] the relative importance of particular environmental variables for a species may vary according to the geographic and biotic contexts".

A keystone species is a species that has a disproportionately large effect on its natural environment relative to its abundance. The concept was introduced in 1969 by the zoologist Robert T. Paine. Keystone species play a critical role in maintaining the structure of an ecological community, affecting many other organisms in an ecosystem and helping to determine the types and numbers of various other species in the community. Without keystone species, the ecosystem would be dramatically different or cease to exist altogether. Some keystone species, such as the wolf and lion are also apex predators.

In ecology, a biological interaction is the effect that a pair of organisms living together in a community have on each other. They can be either of the same species, or of different species. These effects may be short-term, or long-term, both often strongly influence the adaptation and evolution of the species involved. Biological interactions range from mutualism, beneficial to both partners, to competition, harmful to both partners. Interactions can be direct when physical contact is established or indirect, through intermediaries such as shared resources, territories, ecological services, metabolic waste, toxins or growth inhibitors. This type of relationship can be shown by net effect based on individual effects on both organisms arising out of relationship.
This glossary of ecology is a list of definitions of terms and concepts in ecology and related fields. For more specific definitions from other glossaries related to ecology, see Glossary of biology, Glossary of evolutionary biology, and Glossary of environmental science.

In ecology, the competitive exclusion principle, sometimes referred to as Gause's law, is a proposition that two species which compete for the same limited resource cannot coexist at constant population values. When one species has even the slightest advantage over another, the one with the advantage will dominate in the long term. This leads either to the extinction of the weaker competitor or to an evolutionary or behavioral shift toward a different ecological niche. The principle has been paraphrased in the maxim "complete competitors cannot coexist".

An ecosystem engineer is any species that creates, significantly modifies, maintains or destroys a habitat. These organisms can have a large impact on species richness and landscape-level heterogeneity of an area. As a result, ecosystem engineers are important for maintaining the health and stability of the environment they are living in. Since all organisms impact the environment they live in one way or another, it has been proposed that the term "ecosystem engineers" be used only for keystone species whose behavior very strongly affects other organisms.

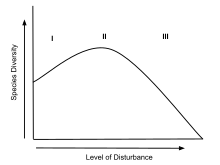
In ecology, a disturbance is a temporary change in environmental conditions that causes a pronounced change in an ecosystem. Disturbances often act quickly and with great effect, to alter the physical structure or arrangement of biotic and abiotic elements. A disturbance can also occur over a long period of time and can impact the biodiversity within an ecosystem.

Species richness, or biodiversity, increases from the poles to the tropics for a wide variety of terrestrial and marine organisms, often referred to as the latitudinal diversity gradient. The latitudinal diversity gradient is one of the most widely recognized patterns in ecology. It has been observed to varying degrees in Earth's past. A parallel trend has been found with elevation, though this is less well-studied.
Ecological facilitation or probiosis describes species interactions that benefit at least one of the participants and cause harm to neither. Facilitations can be categorized as mutualisms, in which both species benefit, or commensalisms, in which one species benefits and the other is unaffected. This article addresses both the mechanisms of facilitation and the increasing information available concerning the impacts of facilitation on community ecology.

A treefall gap is a distinguishable hole in the canopy of a forest with vertical sides extending through all levels down to an average height of 2 m (6.6 ft) above ground. These holes occur as result of a fallen tree or large limb. The ecologist who developed this definition used two meters because he believed that "a regrowth height of 2 m was sufficient" for a gap to be considered closed, but not all scientists agree. For example, Runkle believed that regrowth should be 10–20 m (33–66 ft) above the ground. Alternatively, a treefall gap is "the smallest gap [that must] be readily distinguishable amid the complexity of forest structure."

Competition is an interaction between organisms or species in which both require a resource that is in limited supply. Competition lowers the fitness of both organisms involved since the presence of one of the organisms always reduces the amount of the resource available to the other.

Interspecific competition, in ecology, is a form of competition in which individuals of different species compete for the same resources in an ecosystem. This can be contrasted with mutualism, a type of symbiosis. Competition between members of the same species is called intraspecific competition.
Trophic cascades are powerful indirect interactions that can control entire ecosystems, occurring when a trophic level in a food web is suppressed. For example, a top-down cascade will occur if predators are effective enough in predation to reduce the abundance, or alter the behavior of their prey, thereby releasing the next lower trophic level from predation.

In ecology, a community is a group or association of populations of two or more different species occupying the same geographical area at the same time, also known as a biocoenosis, biotic community, biological community, ecological community, or life assemblage. The term community has a variety of uses. In its simplest form it refers to groups of organisms in a specific place or time, for example, "the fish community of Lake Ontario before industrialization".
In ecology, the theory of alternative stable states predicts that ecosystems can exist under multiple "states". These alternative states are non-transitory and therefore considered stable over ecologically-relevant timescales. Ecosystems may transition from one stable state to another, in what is known as a state shift, when perturbed. Due to ecological feedbacks, ecosystems display resistance to state shifts and therefore tend to remain in one state unless perturbations are large enough. Multiple states may persist under equal environmental conditions, a phenomenon known as hysteresis. Alternative stable state theory suggests that discrete states are separated by ecological thresholds, in contrast to ecosystems which change smoothly and continuously along an environmental gradient.
In ecology, a priority effect refers to the impact that a particular species can have on community development as a result of its prior arrival at a site. There are two basic types of priority effects: inhibitory and facilitative. An inhibitory priority effect occurs when a species that arrives first at a site negatively affects a species that arrives later by reducing the availability of space or resources. In contrast, a facilitative priority effect occurs when a species that arrives first at a site alters abiotic or biotic conditions in ways that positively affect a species that arrives later. Inhibitory priority effects have been documented more frequently than facilitative priority effects. Studies indicate that both abiotic and biotic factors can affect the strength of priority effects. Priority effects are a central and pervasive element of ecological community development that have significant implications for natural systems and ecological restoration efforts.
The Janzen–Connell hypothesis is a well-known hypothesis for the maintenance of high species biodiversity in the tropics. It was published independently in the early 1970s by Daniel Janzen, who focused on tropical trees, and Joseph Connell who discussed trees and marine invertebrates. According to their hypothesis, host-specific herbivores, pathogens, or other natural enemies make the areas near a parent tree inhospitable for the survival of seeds or seedlings. These natural enemies are referred to as 'distance-responsive predators' if they kill seeds or seedlings near the parent tree, or 'density-dependent predators' if they kill seeds or seedlings where they are most abundant. Such predators can prevent any one species from dominating the landscape, because if that species is too common, there will be few safe places for its seedlings to survive. Both Janzen and Connell originally proposed that for natural enemies to increase local diversity, they must be host-specific and relatively immobile, such that they disproportionately reduce the density of the more locally common tree species. This prevents any one species from becoming dominant and excluding other species through competition, allowing more species to coexist in small areas. This can be classified as a stabilizing mechanism.

Intraguild predation, or IGP, is the killing and sometimes eating of a potential competitor of a different species. This interaction represents a combination of predation and competition, because both species rely on the same prey resources and also benefit from preying upon one another. Intraguild predation is common in nature and can be asymmetrical, in which one species feeds upon the other, or symmetrical, in which both species prey upon each other. Because the dominant intraguild predator gains the dual benefits of feeding and eliminating a potential competitor, IGP interactions can have considerable effects on the structure of ecological communities.
Joseph Hurd Connell FAA was an American ecologist. He earned his MA degree in zoology at the University of California, Berkeley and his PhD at Glasgow University. Connell's first research paper examined the effects of interspecific competition and predation on populations of a barnacle species on the rocky shores of Scotland. According to Connell, this classic paper is often cited because it addressed ecological topics that previously had been given minor roles. Together, with a subsequent barnacle study on the influence of competition and desiccation, these two influential papers have laid the foundation for future research and the findings continue to have relevance to current ecology. His early work earned him a Guggenheim fellowship in 1962 and the George Mercer Award in 1963.

A facilitation cascade is a sequence of ecological interactions that occur when a species benefits a second species that in turn has a positive effect on a third species. These facilitative interactions can take the form of amelioration of environmental stress and/or provision of refuge from predation. Autogenic ecosystem engineering species, structural species, habitat-forming species, and foundation species are associated with the most commonly recognized examples of facilitation cascades, sometimes referred to as a habitat cascades. Facilitation generally is a much broader concept that includes all forms of positive interactions including pollination, seed dispersal, and co-evolved commensalism and mutualistic relationships, such as between cnidarian hosts and symbiodinium in corals, and between algae and fungi in lichens. As such, facilitation cascades are widespread through all of the earth's major biomes with consistently positive effects on the abundance and biodiversity of associated organisms.
![Disturbed vegetation due to milpa farming. Cayo District, Belize. [Macrae 2008]. IDH example1.jpg](http://upload.wikimedia.org/wikipedia/commons/thumb/5/5e/IDH_example1.jpg/220px-IDH_example1.jpg)
![(Disturbed vegetation due to milpa farming, Contreras Valley, Cayo District, Belize [Macrae 2010]. IDH example2.jpg](http://upload.wikimedia.org/wikipedia/commons/thumb/a/ad/IDH_example2.jpg/220px-IDH_example2.jpg)
![Disturbance due to tree fall, Gainesville, Florida [Daniel 2012]. IDH example3.jpg](http://upload.wikimedia.org/wikipedia/commons/3/35/IDH_example3.jpg)