
The thymus is a specialized primary lymphoid organ of the immune system. Within the thymus, thymus cell lymphocytes or T cells mature. T cells are critical to the adaptive immune system, where the body adapts to specific foreign invaders. The thymus is located in the upper front part of the chest, in the anterior superior mediastinum, behind the sternum, and in front of the heart. It is made up of two lobes, each consisting of a central medulla and an outer cortex, surrounded by a capsule.

T cells are one of the important types of white blood cells of the immune system and play a central role in the adaptive immune response. T cells can be distinguished from other lymphocytes by the presence of a T-cell receptor (TCR) on their cell surface.
The regulatory T cells (Tregs or Treg cells), formerly known as suppressor T cells, are a subpopulation of T cells that modulate the immune system, maintain tolerance to self-antigens, and prevent autoimmune disease. Treg cells are immunosuppressive and generally suppress or downregulate induction and proliferation of effector T cells. Treg cells express the biomarkers CD4, FOXP3, and CD25 and are thought to be derived from the same lineage as naïve CD4+ cells. Because effector T cells also express CD4 and CD25, Treg cells are very difficult to effectively discern from effector CD4+, making them difficult to study. Research has found that the cytokine transforming growth factor beta (TGF-β) is essential for Treg cells to differentiate from naïve CD4+ cells and is important in maintaining Treg cell homeostasis.
Cross-presentation is the ability of certain professional antigen-presenting cells (mostly dendritic cells) to take up, process and present extracellular antigens with MHC class I molecules to CD8 T cells (cytotoxic T cells). Cross-priming, the result of this process, describes the stimulation of naive cytotoxic CD8+ T cells into activated cytotoxic CD8+ T cells. This process is necessary for immunity against most tumors and against viruses that infect dendritic cells and sabotage their presentation of virus antigens. Cross presentation is also required for the induction of cytotoxic immunity by vaccination with protein antigens, for example, tumour vaccination.
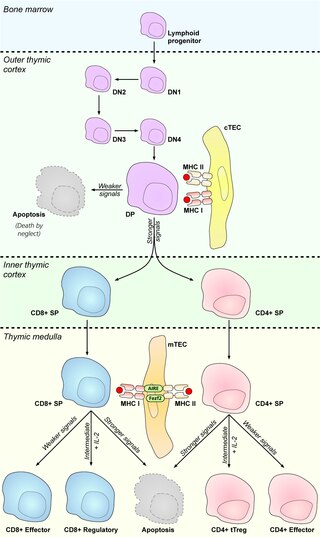
In immunology, central tolerance is the process of eliminating any developing T or B lymphocytes that are autoreactive, i.e. reactive to the body itself. Through elimination of autoreactive lymphocytes, tolerance ensures that the immune system does not attack self peptides. Lymphocyte maturation occurs in primary lymphoid organs such as the bone marrow and the thymus. In mammals, B cells mature in the bone marrow and T cells mature in the thymus.
Immune tolerance, or immunological tolerance, or immunotolerance, is a state of unresponsiveness of the immune system to substances or tissues that would otherwise have the capacity to elicit an immune response in a given organism. It is induced by prior exposure to that specific antigen and contrasts with conventional immune-mediated elimination of foreign antigens. Tolerance is classified into central tolerance or peripheral tolerance depending on where the state is originally induced—in the thymus and bone marrow (central) or in other tissues and lymph nodes (peripheral). The mechanisms by which these forms of tolerance are established are distinct, but the resulting effect is similar.
A thymocyte is an immune cell present in the thymus, before it undergoes transformation into a T cell. Thymocytes are produced as stem cells in the bone marrow and reach the thymus via the blood.

Hassall's corpuscles (or thymic corpuscles (bodies)) are structures found in the medulla of the human thymus, formed from eosinophilic type VI epithelial reticular cells arranged concentrically. These concentric corpuscles are composed of a central mass, consisting of one or more granular cells, and of a capsule formed of epithelioid cells. They vary in size with diameters from 20 to more than 100μm, and tend to grow larger with age. They can be spherical or ovoid and their epithelial cells contain keratohyalin and bundles of cytoplasmic fibres. Later studies indicate that Hassall's corpuscles differentiate from medullary thymic epithelial cells after they lose autoimmune regulator (AIRE) expression. This makes them an example of Thymic mimetic cells. They are named for Arthur Hill Hassall, who discovered them in 1846.

The autoimmune regulator (AIRE) is a protein that in humans is encoded by the AIRE gene. It is a 13kb gene on chromosome 21q22.3 that has 545 amino acids. AIRE is a transcription factor expressed in the medulla of the thymus. It is part of the mechanism which eliminates self-reactive T cells that would cause autoimmune disease. It exposes T cells to normal, healthy proteins from all parts of the body, and T cells that react to those proteins are destroyed.
Self-protein refers to all proteins endogenously produced by DNA-level transcription and translation within an organism of interest. This does not include proteins synthesized due to viral infection, but may include those synthesized by commensal bacteria within the intestines. Proteins that are not created within the body of the organism of interest, but nevertheless enter through the bloodstream, a breach in the skin, or a mucous membrane, may be designated as “non-self” and subsequently targeted and attacked by the immune system. Tolerance to self-protein is crucial for overall wellbeing; when the body erroneously identifies self-proteins as “non-self”, the subsequent immune response against endogenous proteins may lead to the development of an autoimmune disease.
In immunology, peripheral tolerance is the second branch of immunological tolerance, after central tolerance. It takes place in the immune periphery. Its main purpose is to ensure that self-reactive T and B cells which escaped central tolerance do not cause autoimmune disease. Peripheral tolerance prevents immune response to harmless food antigens and allergens, too.

DNA-binding protein Ikaros also known as Ikaros family zinc finger protein 1 is a protein that in humans is encoded by the IKZF1 gene.
In immunology, clonal deletion is the removal through apoptosis of B cells and T cells that have expressed receptors for self before developing into fully immunocompetent lymphocytes. This prevents recognition and destruction of self host cells, making it a type of negative selection or central tolerance. Central tolerance prevents B and T lymphocytes from reacting to self. Thus, clonal deletion can help protect individuals against autoimmunity. Clonal deletion is thought to be the most common type of negative selection. It is one method of immune tolerance.
Thymic nurse cells (TNCs) are large epithelial cells found in the cortex of the thymus and also in cortico-medullary junction. They have their own nucleus and are known to internalize thymocytes through extensions of plasma membrane. The cell surfaces of TNCs and their cytoplasmic vacuoles express MHC Class I and MHC Class II antigens. The interaction of these antigens with the developing thymocytes determines whether the thymocytes undergo positive or negative selection.
Antigen transfer in the thymus is the transmission of self-antigens between thymic antigen-presenting cells which contributes to the establishment of T cell central tolerance.
Cortical thymic epithelial cells (cTECs) form unique parenchyma cell population of the thymus which critically contribute to the development of T cells.
Thymic epithelial cells (TECs) are specialized cells with high degree of anatomic, phenotypic and functional heterogeneity that are located in the outer layer (epithelium) of the thymic stroma. The thymus, as a primary lymphoid organ, mediates T cell development and maturation. The thymic microenvironment is established by TEC network filled with thymocytes in different developing stages. TECs and thymocytes are the most important components in the thymus, that are necessary for production of functionally competent T lymphocytes and self tolerance. Dysfunction of TECs causes several immunodeficiencies and autoimmune diseases.
Promiscuous gene expression (PGE), formerly referred to as ectopic expression, is a process specific to the thymus that plays a pivotal role in the establishment of central tolerance. This phenomenon enables generation of self-antigens, so called tissue-restricted antigens (TRAs), which are in the body expressed only by one or few specific tissues. These antigens are represented for example by insulin from the pancreas or defensins from the gastrointestinal tract. Antigen-presenting cells (APCs) of the thymus, namely medullary thymic epithelial cells (mTECs), dendritic cells (DCs) and B cells are capable to present peptides derived from TRAs to developing T cells and hereby test, whether their T cell receptors (TCRs) engage self entities and therefore their occurrence in the body can potentially lead to the development of autoimmune disease. In that case, thymic APCs either induce apoptosis in these autoreactive T cells or they deviate them to become T regulatory cells, which suppress self-reactive T cells in the body that escaped negative selection in the thymus. Thus, PGE is crucial for tissues protection against autoimmunity.
Thymus stromal cells are subsets of specialized cells located in different areas of the thymus. They include all non-T-lineage cells, such as thymic epithelial cells (TECs), endothelial cells, mesenchymal cells, dendritic cells, and B lymphocytes, and provide signals essential for thymocyte development and the homeostasis of the thymic stroma.
Thymic mimetic cells are a heterogeneous population of cells located in the thymus that exhibit phenotypes of a wide variety of differentiated peripheral cells. They arise from medullary thymic epithelial cells (mTECs) and also function in negative selection of self-reactive T cells.
