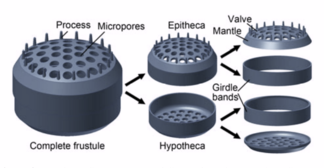
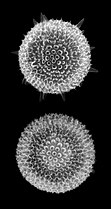
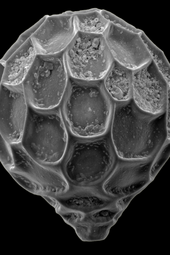
A diatom is any member of a large group comprising several genera of algae, specifically microalgae, found in the oceans, waterways and soils of the world. Living diatoms make up a significant portion of the Earth's biomass: they generate about 20 to 50 percent of the oxygen produced on the planet each year, take in over 6.7 billion tonnes of silicon each year from the waters in which they live, and constitute nearly half of the organic material found in the oceans. The shells of dead diatoms can reach as much as a half-mile deep on the ocean floor, and the entire Amazon basin is fertilized annually by 27 million tons of diatom shell dust transported by transatlantic winds from the African Sahara, much of it from the Bodélé Depression, which was once made up of a system of fresh-water lakes.

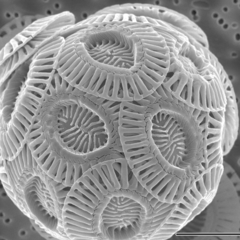
Coccolithophores, or coccolithophorids, are single-celled organisms which are part of the phytoplankton, the autotrophic (self-feeding) component of the plankton community. They form a group of about 200 species, and belong either to the kingdom Protista, according to Robert Whittaker's five-kingdom system, or clade Hacrobia, according to a newer biological classification system. Within the Hacrobia, the coccolithophores are in the phylum or division Haptophyta, class Prymnesiophyceae. Coccolithophores are almost exclusively marine, are photosynthetic, and exist in large numbers throughout the sunlight zone of the ocean.
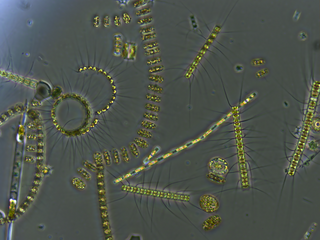
Zooplankton are the animal component of the planktonic community, having to consume other organisms to thrive. Plankton are aquatic organisms that are unable to swim effectively against currents. Consequently, they drift or are carried along by currents in the ocean, or by currents in seas, lakes or rivers.

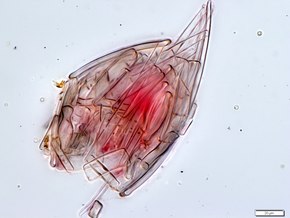
The Radiolaria, also called Radiozoa, are protozoa of diameter 0.1–0.2 mm that produce intricate mineral skeletons, typically with a central capsule dividing the cell into the inner and outer portions of endoplasm and ectoplasm. The elaborate mineral skeleton is usually made of silica. They are found as zooplankton throughout the global ocean. As zooplankton, radiolarians are primarily heterotrophic, but many have photosynthetic endosymbionts and are, therefore, considered mixotrophs. The skeletal remains of some types of radiolarians make up a large part of the cover of the ocean floor as siliceous ooze. Due to their rapid change as species and intricate skeletons, radiolarians represent an important diagnostic fossil found from the Cambrian onwards.
Coccoliths are individual plates or scales of calcium carbonate formed by coccolithophores and cover the cell surface arranged in the form of a spherical shell, called a coccosphere.

Micropaleontology is the branch of paleontology (palaeontology) that studies microfossils, or fossils that require the use of a microscope to see the organism, its morphology and its characteristic details.
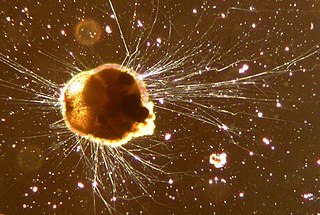
Foraminifera are single-celled organisms, members of a phylum or class of Cercozoan protists characterized by streaming granular ectoplasm for catching food and other uses; and commonly an external shell of diverse forms and materials. Tests of chitin are believed to be the most primitive type. Most foraminifera are marine, the majority of which live on or within the seafloor sediment, while a smaller number float in the water column at various depths, which belong to the suborder Globigerinina. Fewer are known from freshwater or brackish conditions, and some very few (nonaquatic) soil species have been identified through molecular analysis of small subunit ribosomal DNA.

A microfossil is a fossil that is generally between 0.001 mm and 1 mm in size, the visual study of which requires the use of light or electron microscopy. A fossil which can be studied with the naked eye or low-powered magnification, such as a hand lens, is referred to as a macrofossil.
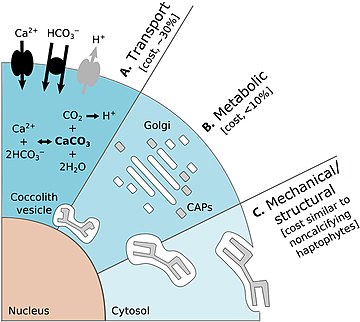
Biomineralization, also written biomineralisation, is the process by which living organisms produce minerals, often resulting in hardened or stiffened mineralized tissues. It is an extremely widespread phenomenon: all six taxonomic kingdoms contain members that are able to form minerals, and over 60 different minerals have been identified in organisms. Examples include silicates in algae and diatoms, carbonates in invertebrates, and calcium phosphates and carbonates in vertebrates. These minerals often form structural features such as sea shells and the bone in mammals and birds.
Biogenic silica (bSi), also referred to as opal, biogenic opal, or amorphous opaline silica, forms one of the most widespread biogenic minerals. For example, microscopic particles of silica called phytoliths can be found in grasses and other plants.

In biology, a test is the hard shell of some spherical marine animals and protists, notably sea urchins and microorganisms such as testate foraminiferans, radiolarians, and testate amoebae. The term is also applied to the covering of scale insects. The related Latin term testa is used for the hard seed coat of plant seeds.

Marine sediment, or ocean sediment, or seafloor sediment, are deposits of insoluble particles that have accumulated on the seafloor. These particles either have their origins in soil and rocks and have been transported from the land to the sea, mainly by rivers but also by dust carried by wind and by the flow of glaciers into the sea, or they are biogenic deposits from marine organisms or from chemical precipitation in seawater, as well as from underwater volcanoes and meteorite debris.

Siliceous ooze is a type of biogenic pelagic sediment located on the deep ocean floor. Siliceous oozes are the least common of the deep sea sediments, and make up approximately 15% of the ocean floor. Oozes are defined as sediments which contain at least 30% skeletal remains of pelagic microorganisms. Siliceous oozes are largely composed of the silica based skeletons of microscopic marine organisms such as diatoms and radiolarians. Other components of siliceous oozes near continental margins may include terrestrially derived silica particles and sponge spicules. Siliceous oozes are composed of skeletons made from opal silica SiO2·nH2O, as opposed to calcareous oozes, which are made from skeletons of calcium carbonate (CaCO3·nH2O) organisms (i.e. coccolithophores). Silica (Si) is a bioessential element and is efficiently recycled in the marine environment through the silica cycle. Distance from land masses, water depth and ocean fertility are all factors that affect the opal silica content in seawater and the presence of siliceous oozes.

Marine microorganisms are defined by their habitat as microorganisms living in a marine environment, that is, in the saltwater of a sea or ocean or the brackish water of a coastal estuary. A microorganism is any microscopic living organism or virus, that is too small to see with the unaided human eye without magnification. Microorganisms are very diverse. They can be single-celled or multicellular and include bacteria, archaea, viruses and most protozoa, as well as some fungi, algae, and animals, such as rotifers and copepods. Many macroscopic animals and plants have microscopic juvenile stages. Some microbiologists also classify viruses as microorganisms, but others consider these as non-living.
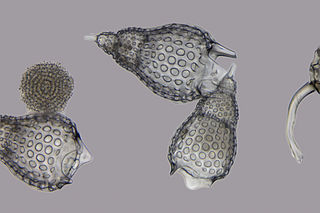
Nassellaria is an order of Rhizaria belonging to the class Radiolaria. The organisms of this order are characterized by a skeleton cross link with a cone or ring.
Marine biogenic calcification is the production of calcium carbonate by organisms in the global ocean.

Marine protists are defined by their habitat as protists that live in marine environments, that is, in the saltwater of seas or oceans or the brackish water of coastal estuaries. Life originated as marine single-celled prokaryotes and later evolved into more complex eukaryotes. Eukaryotes are the more developed life forms known as plants, animals, fungi and protists. Protists are the eukaryotes that cannot be classified as plants, fungi or animals. They are mostly single-celled and microscopic. The term protist came into use historically as a term of convenience for eukaryotes that cannot be strictly classified as plants, animals or fungi. They are not a part of modern cladistics because they are paraphyletic.

Foraminiferal tests are the tests of Foraminifera.
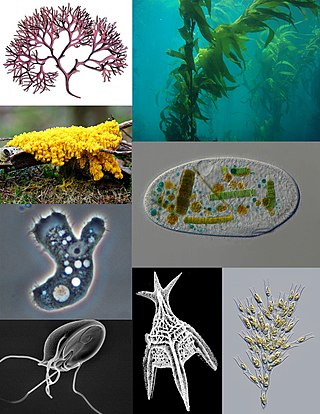
A protist is any eukaryotic organism that is not an animal, plant, or fungus. While it is likely that protists share a common ancestor, the last eukaryotic common ancestor, the exclusion of other eukaryotes means that protists do not form a natural group, or clade. Therefore, some protists may be more closely related to animals, plants, or fungi than they are to other protists. However, like algae, invertebrates and protozoans, the grouping is used for convenience.
Global paleoclimate indicators are the proxies sensitive to global paleoclimatic environment changes. They are mostly derived from marine sediments. Paleoclimate indicators derived from terrestrial sediments, on the other hand, are commonly influenced by local tectonic movements and paleogeographic variations. Factors governing the Earth's climate system include plate tectonics, which controls the configuration of continents, the interplay between the atmosphere and the ocean, and the Earth's orbital characteristics. Global paleoclimate indicators are established based on the information extracted from the analyses of geologic materials, including biological, geochemical and mineralogical data preserved in marine sediments. Indicators are generally grouped into three categories; paleontological, geochemical and lithological.