
Sexual selection is a mode of natural selection in which members of one biological sex choose mates of the other sex to mate with, and compete with members of the same sex for access to members of the opposite sex. These two forms of selection mean that some individuals have greater reproductive success than others within a population, for example because they are more attractive or prefer more attractive partners to produce offspring. Successful males benefit from frequent mating and monopolizing access to one or more fertile females. Females can maximise the return on the energy they invest in reproduction by selecting and mating with the best males.

Mealworms are the larval form of the yellow mealworm beetle, Tenebrio molitor, a species of darkling beetle. Like all holometabolic insects, they go through four life stages: egg, larva, pupa, and adult. Larvae typically measure about 2.5 centimetres (0.98 in) or more, whereas adults are generally 1.25 to 1.8 centimetres in length.

Sperm competition is the competitive process between spermatozoa of two or more different males to fertilize the same egg during sexual reproduction. Competition can occur when females have multiple potential mating partners. Greater choice and variety of mates increases a female's chance to produce more viable offspring. However, multiple mates for a female means each individual male has decreased chances of producing offspring. Sperm competition is an evolutionary pressure on males, and has led to the development of adaptations to increase male's chance of reproductive success. Sperm competition results in a sexual conflict between males and females. Males have evolved several defensive tactics including: mate-guarding, mating plugs, and releasing toxic seminal substances to reduce female re-mating tendencies to cope with sperm competition. Offensive tactics of sperm competition involve direct interference by one male on the reproductive success of another male, for instance by mate guarding or by physically removing another male's sperm prior to mating with a female. For an example, see Gryllus bimaculatus.

Parental investment, in evolutionary biology and evolutionary psychology, is any parental expenditure that benefits offspring. Parental investment may be performed by both males and females, females alone or males alone. Care can be provided at any stage of the offspring's life, from pre-natal to post-natal.
Philopatry is the tendency of an organism to stay in or habitually return to a particular area. The causes of philopatry are numerous, but natal philopatry, where animals return to their birthplace to breed, may be the most common. The term derives from the Greek roots philo, "liking, loving" and patra, "fatherland", although in recent years the term has been applied to more than just the animal's birthplace. Recent usage refers to animals returning to the same area to breed despite not being born there, and migratory species that demonstrate site fidelity: reusing stopovers, staging points, and wintering grounds.
Life history theory (LHT) is an analytical framework designed to study the diversity of life history strategies used by different organisms throughout the world, as well as the causes and results of the variation in their life cycles. It is a theory of biological evolution that seeks to explain aspects of organisms' anatomy and behavior by reference to the way that their life histories—including their reproductive development and behaviors, post-reproductive behaviors, and lifespan —have been shaped by natural selection. A life history strategy is the "age- and stage-specific patterns" and timing of events that make up an organism's life, such as birth, weaning, maturation, death, etc. These events, notably juvenile development, age of sexual maturity, first reproduction, number of offspring and level of parental investment, senescence and death, depend on the physical and ecological environment of the organism.
Monogamous pairing in animals refers to the natural history of mating systems in which species pair bond to raise offspring. This is associated, usually implicitly, with sexual monogamy.
Cooperative breeding is a social system characterized by alloparental care: offspring receive care not only from their parents, but also from additional group members, often called helpers. Cooperative breeding encompasses a wide variety of group structures, from a breeding pair with helpers that are offspring from a previous season, to groups with multiple breeding males and females (polygynandry) and helpers that are the adult offspring of some but not all of the breeders in the group, to groups in which helpers sometimes achieve co-breeding status by producing their own offspring as part of the group's brood. Cooperative breeding occurs across taxonomic groups including birds, mammals, fish, and insects.

Sexual conflict or sexual antagonism occurs when the two sexes have conflicting optimal fitness strategies concerning reproduction, particularly over the mode and frequency of mating, potentially leading to an evolutionary arms race between males and females. In one example, males may benefit from multiple matings, while multiple matings may harm or endanger females, due to the anatomical differences of that species. Sexual conflict underlies the evolutionary distinction between male and female.

Mate choice is one of the primary mechanisms under which evolution can occur. It is characterized by a "selective response by animals to particular stimuli" which can be observed as behavior. In other words, before an animal engages with a potential mate, they first evaluate various aspects of that mate which are indicative of quality—such as the resources or phenotypes they have—and evaluate whether or not those particular trait(s) are somehow beneficial to them. The evaluation will then incur a response of some sort.

The sexy son hypothesis in evolutionary biology and sexual selection, proposed by Patrick J. Weatherhead and Raleigh J. Robertson of Queen's University in Kingston, Ontario in 1979, states that a female's ideal mate choice among potential mates is one whose genes will produce males with the best chance of reproductive success. This implies that other benefits the father can offer the mother or offspring are less relevant than they may appear, including his capacity as a parental caregiver, territory and any nuptial gifts. Fisher's principle means that the sex ratio is always near 1:1 between males and females, yet what matters most are her "sexy sons'" future breeding successes, more likely if they have a promiscuous father, in creating large numbers of offspring carrying copies of her genes. This sexual selection hypothesis has been researched in species such as the European pied flycatcher.
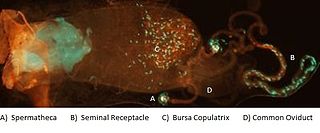
Female sperm storage is a biological process and often a type of sexual selection in which sperm cells transferred to a female during mating are temporarily retained within a specific part of the reproductive tract before the oocyte, or egg, is fertilized. This process takes place in some species of animals, but not in humans. The site of storage is variable among different animal taxa and ranges from structures that appear to function solely for sperm retention, such as insect spermatheca and bird sperm storage tubules, to more general regions of the reproductive tract enriched with receptors to which sperm associate before fertilization, such as the caudal portion of the cow oviduct containing sperm-associating annexins. Female sperm storage is an integral stage in the reproductive process for many animals with internal fertilization. It has several documented biological functions including:

Eusociality evolved repeatedly in different orders of animals, notably termites and the Hymenoptera. This 'true sociality' in animals, in which sterile individuals work to further the reproductive success of others, is found in termites, ambrosia beetles, gall-dwelling aphids, thrips, marine sponge-dwelling shrimp, naked mole-rats, and many genera in the insect order Hymenoptera. The fact that eusociality has evolved so often in the Hymenoptera, but remains rare throughout the rest of the animal kingdom, has made its evolution a topic of debate among evolutionary biologists. Eusocial organisms at first appear to behave in stark contrast with simple interpretations of Darwinian evolution: passing on one's genes to the next generation, or fitness, is a central idea in evolutionary biology.
A biological ornament is a characteristic of an animal that appears to serve a decorative function rather than a utilitarian function. Many are secondary sexual characteristics, and others appear on young birds during the period when they are dependent on being fed by their parents. Ornaments are used in displays to attract mates, which may lead to the evolutionary process known as sexual selection. An animal may shake, lengthen, or spread out its ornament in order to get the attention of the opposite sex, which will in turn choose the most attractive one with which to mate. Ornaments are most often observed in males, and choosing an extravagantly ornamented male benefits females as the genes that produce the ornament will be passed on to her offspring, increasing their own reproductive fitness. As Ronald Fisher noted, the male offspring will inherit the ornament while the female offspring will inherit the preference for said ornament, which can lead to a positive feedback loop known as a Fisherian runaway. These structures serve as cues to animal sexual behaviour, that is, they are sensory signals that affect mating responses. Therefore, ornamental traits are often selected by mate choice.
Interlocus sexual conflict is a type of sexual conflict that occurs through the interaction of a set of antagonistic alleles at two or more different loci, or the location of a gene on a chromosome, in males and females, resulting in the deviation of either or both sexes from the fitness optima for the traits. A co-evolutionary arms race is established between the sexes in which either sex evolves a set of antagonistic adaptations that is detrimental to the fitness of the other sex. The potential for reproductive success in one organism is strengthened while the fitness of the opposite sex is weakened. Interlocus sexual conflict can arise due to aspects of male–female interactions such as mating frequency, fertilization, relative parental effort, female remating behavior, and female reproductive rate.
The theoretical foundations of evolutionary psychology are the general and specific scientific theories that explain the ultimate origins of psychological traits in terms of evolution. These theories originated with Charles Darwin's work, including his speculations about the evolutionary origins of social instincts in humans. Modern evolutionary psychology, however, is possible only because of advances in evolutionary theory in the 20th century.
Filial cannibalism occurs when an adult individual of a species consumes all or part of the young of its own species or immediate offspring. Filial cannibalism occurs in many species ranging from mammals to insects, and is especially prevalent in various types of fish species with males that engage in egg guardianship. The exact evolutionary purpose of the practice in those species is unclear and debated among zoologists, though there is consensus that it may have, or may have had at some point in species' evolutionary history, certain evolutionary and ecological implications.
Inbreeding avoidance, or the inbreeding avoidance hypothesis, is a concept in evolutionary biology that refers to the prevention of the deleterious effects of inbreeding. Animals only rarely exhibit inbreeding avoidance. The inbreeding avoidance hypothesis posits that certain mechanisms develop within a species, or within a given population of a species, as a result of assortative mating and natural and sexual selection, in order to prevent breeding among related individuals. Although inbreeding may impose certain evolutionary costs, inbreeding avoidance, which limits the number of potential mates for a given individual, can inflict opportunity costs. Therefore, a balance exists between inbreeding and inbreeding avoidance. This balance determines whether inbreeding mechanisms develop and the specific nature of such mechanisms.
Human reproductive ecology is a subfield in evolutionary biology that is concerned with human reproductive processes and responses to ecological variables. It is based in the natural and social sciences, and is based on theory and models deriving from human and animal biology, evolutionary theory, and ecology. It is associated with fields such as evolutionary anthropology and seeks to explain human reproductive variation and adaptations. The theoretical orientation of reproductive ecology applies the theory of natural selection to reproductive behaviors, and has also been referred to as the evolutionary ecology of human reproduction.

Gnatocerus cornutus, the broadhorned flour beetle, is a common species of beetle in the Tenebrionidae family. It is distributed worldwide and is commonly found in warm, tropical regions. The total development time is about 47 days. G. cornutus is predominately found in mills as a storage pest and has a wide distribution.
