
Peafowl is a common name for two bird species in the genera Pavo and Afropavo within the tribe Pavonini of the family Phasianidae. Male peafowl are referred to as peacocks, and female peafowl are referred to as peahens, although peafowl of either sex are often referred to colloquially as "peacocks".

Sexual dimorphism is the condition where sexes of the same species exhibit different morphological characteristics, particularly characteristics not directly involved in reproduction. The condition occurs in most dioecious species, which consist of most animals and some plants. Differences may include secondary sex characteristics, size, weight, color, markings, or behavioral or cognitive traits. Male-male reproductive competition has evolved a diverse array of sexually dimorphic traits. Aggressive utility traits such as "battle" teeth and blunt heads reinforced as battering rams are used as weapons in aggressive interactions between rivals. Passive displays such as ornamental feathering or song-calling have also evolved mainly through sexual selection. These differences may be subtle or exaggerated and may be subjected to sexual selection and natural selection. The opposite of dimorphism is monomorphism, when both biological sexes are phenotypically indistinguishable from each other.

A lek is an aggregation of male animals gathered to engage in competitive displays and courtship rituals, known as lekking, to entice visiting females which are surveying prospective partners with which to mate. A lek can also indicate an available plot of space able to be utilized by displaying males to defend their own share of territory for the breeding season. A lekking species is characterised by male displays, strong female mate choice, and the conferring of indirect benefits to males and reduced costs to females. Although most prevalent among birds such as black grouse, lekking is also found in a wide range of vertebrates including some bony fish, amphibians, reptiles, and mammals, and arthropods including crustaceans and insects.

Behavioral ecology, also spelled behavioural ecology, is the study of the evolutionary basis for animal behavior due to ecological pressures. Behavioral ecology emerged from ethology after Niko Tinbergen outlined four questions to address when studying animal behaviors: What are the proximate causes, ontogeny, survival value, and phylogeny of a behavior?

Fisherian runaway or runaway selection is a sexual selection mechanism proposed by the mathematical biologist Ronald Fisher in the early 20th century, to account for the evolution of ostentatious male ornamentation by persistent, directional female choice. An example is the colourful and elaborate peacock plumage compared to the relatively subdued peahen plumage; the costly ornaments, notably the bird's extremely long tail, appear to be incompatible with natural selection. Fisherian runaway can be postulated to include sexually dimorphic phenotypic traits such as behavior expressed by a particular sex.
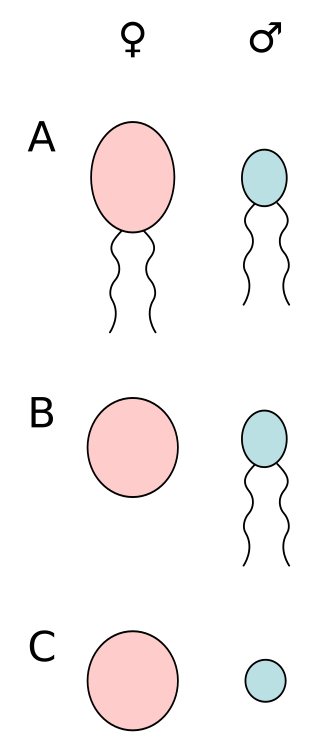
Anisogamy is a form of sexual reproduction that involves the union or fusion of two gametes that differ in size and/or form. The smaller gamete is male, a sperm cell, whereas the larger gamete is female, typically an egg cell. Anisogamy is predominant among multicellular organisms. In both plants and animals gamete size difference is the fundamental difference between females and males.

A psychological adaptation is a functional, cognitive or behavioral trait that benefits an organism in its environment. Psychological adaptations fall under the scope of evolved psychological mechanisms (EPMs), however, EPMs refer to a less restricted set. Psychological adaptations include only the functional traits that increase the fitness of an organism, while EPMs refer to any psychological mechanism that developed through the processes of evolution. These additional EPMs are the by-product traits of a species’ evolutionary development, as well as the vestigial traits that no longer benefit the species’ fitness. It can be difficult to tell whether a trait is vestigial or not, so some literature is more lenient and refers to vestigial traits as adaptations, even though they may no longer have adaptive functionality. For example, xenophobic attitudes and behaviors, some have claimed, appear to have certain EPM influences relating to disease aversion, however, in many environments these behaviors will have a detrimental effect on a person's fitness. The principles of psychological adaptation rely on Darwin's theory of evolution and are important to the fields of evolutionary psychology, biology, and cognitive science.

Sexual conflict or sexual antagonism occurs when the two sexes have conflicting optimal fitness strategies concerning reproduction, particularly over the mode and frequency of mating, potentially leading to an evolutionary arms race between males and females. In one example, males may benefit from multiple matings, while multiple matings may harm or endanger females, due to the anatomical differences of that species. Sexual conflict underlies the evolutionary distinction between male and female.

Mate choice is one of the primary mechanisms under which evolution can occur. It is characterized by a "selective response by animals to particular stimuli" which can be observed as behavior. In other words, before an animal engages with a potential mate, they first evaluate various aspects of that mate which are indicative of quality—such as the resources or phenotypes they have—and evaluate whether or not those particular trait(s) are somehow beneficial to them. The evaluation will then incur a response of some sort.

The sexy son hypothesis in evolutionary biology and sexual selection, proposed by Patrick J. Weatherhead and Raleigh J. Robertson of Queen's University in Kingston, Ontario in 1979, states that a female's ideal mate choice among potential mates is one whose genes will produce males with the best chance of reproductive success. This implies that other benefits the father can offer the mother or offspring are less relevant than they may appear, including his capacity as a parental caregiver, territory and any nuptial gifts. Fisher's principle means that the sex ratio is always near 1:1 between males and females, yet what matters most are her "sexy sons'" future breeding successes, more likely if they have a promiscuous father, in creating large numbers of offspring carrying copies of her genes. This sexual selection hypothesis has been researched in species such as the European pied flycatcher.

A courtship display is a set of display behaviors in which an animal, usually a male, attempts to attract a mate; the mate exercises choice, so sexual selection acts on the display. These behaviors often include ritualized movement ("dances"), vocalizations, mechanical sound production, or displays of beauty, strength, or agonistic ability.
Sexual selection in humans concerns the concept of sexual selection, introduced by Charles Darwin as an element of his theory of natural selection, as it affects humans. Sexual selection is a biological way one sex chooses a mate for the best reproductive success. Most compete with others of the same sex for the best mate to contribute their genome for future generations. This has shaped human evolution for many years, but reasons why humans choose their mates are not fully understood. Sexual selection is quite different in non-human animals than humans as they feel more of the evolutionary pressures to reproduce and can easily reject a mate. The role of sexual selection in human evolution has not been firmly established although neoteny has been cited as being caused by human sexual selection. It has been suggested that sexual selection played a part in the evolution of the anatomically modern human brain, i.e. the structures responsible for social intelligence underwent positive selection as a sexual ornamentation to be used in courtship rather than for survival itself, and that it has developed in ways outlined by Ronald Fisher in the Fisherian runaway model. Fisher also stated that the development of sexual selection was "more favourable" in humans.
Sexual antagonistic co-evolution is the relationship between males and females where sexual morphology changes over time to counteract the opposite's sex traits to achieve the maximum reproductive success. This has been compared to an arms race between sexes. In many cases, male mating behavior is detrimental to the female's fitness. For example, when insects reproduce by means of traumatic insemination, it is very disadvantageous to the female's health. During mating, males will try to inseminate as many females as possible, however, the more times a female's abdomen is punctured, the less likely she is to survive. Females that possess traits to avoid multiple matings will be more likely to survive, resulting in a change in morphology. In males, genitalia is relatively simple and more likely to vary among generations compared to female genitalia. This results in a new trait that females have to avoid in order to survive.
A biological ornament is a characteristic of an animal that appears to serve a decorative function rather than a utilitarian function. Many are secondary sexual characteristics, and others appear on young birds during the period when they are dependent on being fed by their parents. Ornaments are used in displays to attract mates, which may lead to the evolutionary process known as sexual selection. An animal may shake, lengthen, or spread out its ornament in order to get the attention of the opposite sex, which will in turn choose the most attractive one with which to mate. Ornaments are most often observed in males, and choosing an extravagantly ornamented male benefits females as the genes that produce the ornament will be passed on to her offspring, increasing their own reproductive fitness. As Ronald Fisher noted, the male offspring will inherit the ornament while the female offspring will inherit the preference for said ornament, which can lead to a positive feedback loop known as a Fisherian runaway. These structures serve as cues to animal sexual behaviour, that is, they are sensory signals that affect mating responses. Therefore, ornamental traits are often selected by mate choice.
Interlocus sexual conflict is a type of sexual conflict that occurs through the interaction of a set of antagonistic alleles at two or more different loci, or the location of a gene on a chromosome, in males and females, resulting in the deviation of either or both sexes from the fitness optima for the traits. A co-evolutionary arms race is established between the sexes in which either sex evolves a set of antagonistic adaptations that is detrimental to the fitness of the other sex. The potential for reproductive success in one organism is strengthened while the fitness of the opposite sex is weakened. Interlocus sexual conflict can arise due to aspects of male–female interactions such as mating frequency, fertilization, relative parental effort, female remating behavior, and female reproductive rate.
Evolutionary aesthetics refers to evolutionary psychology theories in which the basic aesthetic preferences of Homo sapiens are argued to have evolved in order to enhance survival and reproductive success.

Sexual selection in birds concerns how birds have evolved a variety of mating behaviors, with the peacock tail being perhaps the most famous example of sexual selection and the Fisherian runaway. Commonly occurring sexual dimorphisms such as size and color differences are energetically costly attributes that signal competitive breeding situations. Many types of avian sexual selection have been identified; intersexual selection, also known as female choice; and intrasexual competition, where individuals of the more abundant sex compete with each other for the privilege to mate. Sexually selected traits often evolve to become more pronounced in competitive breeding situations until the trait begins to limit the individual's fitness. Conflicts between an individual fitness and signaling adaptations ensure that sexually selected ornaments such as plumage coloration and courtship behavior are "honest" traits. Signals must be costly to ensure that only good-quality individuals can present these exaggerated sexual ornaments and behaviors.

Sexual selection in mammals is a process the study of which started with Charles Darwin's observations concerning sexual selection, including sexual selection in humans, and in other mammals, consisting of male–male competition and mate choice that mold the development of future phenotypes in a population for a given species.

Sexual selection in amphibians involves sexual selection processes in amphibians, including frogs, salamanders and newts. Prolonged breeders, the majority of frog species, have breeding seasons at regular intervals where male-male competition occurs with males arriving at the waters edge first in large number and producing a wide range of vocalizations, with variations in depth of calls the speed of calls and other complex behaviours to attract mates. The fittest males will have the deepest croaks and the best territories, with females making their mate choices at least partly based on the males depth of croaking. This has led to sexual dimorphism, with females being larger than males in 90% of species, males in 10% and males fighting for groups of females.

The Evolution of Beauty: How Darwin's Forgotten Theory of Mate Choice Shapes the Animal World—and Us is a 2017 book by the ornithologist and evolutionary biologist Richard O. Prum about the power of aesthetic mate choice, arguing it to be an important independent agent in evolution. Prum indicates that while Charles Darwin made this argument in The Descent of Man, published in 1871, the concept was sidelined and forgotten and the notion of natural selection being the sole driver of evolution took over. As an ornithologist, Prum describes many examples in avian evolution where aesthetics are preeminent. Prum proceeds to apply the principle of aesthetic evolution as an independent force in human evolution.



















