
An autosome is any chromosome that is not a sex chromosome. The members of an autosome pair in a diploid cell have the same morphology, unlike those in allosomal pairs, which may have different structures. The DNA in autosomes is collectively known as atDNA or auDNA.
Sex is the trait that determines whether a sexually reproducing organism produces male or female gametes. During sexual reproduction, a male and a female gamete fuse to form a zygote, which develops into an offspring that inherits traits from each parent. By convention, organisms that produce smaller, more mobile gametes are called male, while organisms that produce larger, non-mobile gametes are called female. An organism that produces both types of gamete is hermaphrodite.
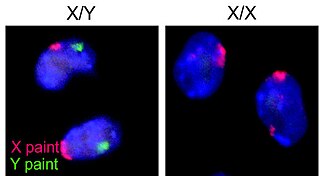
The XY sex-determination system is a sex-determination system used to classify many mammals, including humans, some insects (Drosophila), some snakes, some fish (guppies), and some plants. In this system, the sex of an individual is determined by a pair of sex chromosomes. Females have two of the same kind of sex chromosome (XX), and are called the homogametic sex. Males have two different kinds of sex chromosomes (XY), and are called the heterogametic sex.

A sex-determination system is a biological system that determines the development of sexual characteristics in an organism. Most organisms that create their offspring using sexual reproduction have two common sexes and a few less common intersex variations.

The Y chromosome is one of two sex chromosomes in therian mammals and other organisms. Along with the X chromosome, it is part of the XY sex-determination system, in which the Y is the sex-determining because it is the presence or absence of Y chromosome that determines the male or female sex of offspring produced in sexual reproduction. In mammals, the Y chromosome contains the SRY gene, which triggers development of male gonads. The Y chromosome is passed only from male parents to male offspring.

A pair of homologous chromosomes, or homologs, is a set of one maternal and one paternal chromosome that pair up with each other inside a cell during fertilization. Homologs have the same genes in the same loci, where they provide points along each chromosome that enable a pair of chromosomes to align correctly with each other before separating during meiosis. This is the basis for Mendelian inheritance, which characterizes inheritance patterns of genetic material from an organism to its offspring parent developmental cell at the given time and area.

Haldane's rule is an observation about the early stage of speciation, formulated in 1922 by the British evolutionary biologist J. B. S. Haldane, that states that if — in a species hybrid — only one sex is inviable or sterile, that sex is more likely to be the heterogametic sex. The heterogametic sex is the one with two different sex chromosomes; in therian mammals, for example, this is the male.

Sex linked describes the sex-specific reading patterns of inheritance and presentation when a gene mutation (allele) is present on a sex chromosome (allosome) rather than a non-sex chromosome (autosome). In humans, these are termed X-linked recessive, X-linked dominant and Y-linked. The inheritance and presentation of all three differ depending on the sex of both the parent and the child. This makes them characteristically different from autosomal dominance and recessiveness.

Sex-determining region Y protein (SRY), or testis-determining factor (TDF), is a DNA-binding protein encoded by the SRY gene that is responsible for the initiation of male sex determination in therian mammals. SRY is an intronless sex-determining gene on the Y chromosome. Mutations in this gene lead to a range of disorders of sex development with varying effects on an individual's phenotype and genotype.

Dosage compensation is the process by which organisms equalize the expression of genes between members of different biological sexes. Across species, different sexes are often characterized by different types and numbers of sex chromosomes. In order to neutralize the large difference in gene dosage produced by differing numbers of sex chromosomes among the sexes, various evolutionary branches have acquired various methods to equalize gene expression among the sexes. Because sex chromosomes contain different numbers of genes, different species of organisms have developed different mechanisms to cope with this inequality. Replicating the actual gene is impossible; thus organisms instead equalize the expression from each gene. For example, in humans, female (XX) cells randomly silence the transcription of one X chromosome, and transcribe all information from the other, expressed X chromosome. Thus, human females have the same number of expressed X-linked genes per cell as do human males (XY), both sexes having essentially one X chromosome per cell, from which to transcribe and express genes.
Human genetics is the study of inheritance as it occurs in human beings. Human genetics encompasses a variety of overlapping fields including: classical genetics, cytogenetics, molecular genetics, biochemical genetics, genomics, population genetics, developmental genetics, clinical genetics, and genetic counseling.

Male is the sex of an organism that produces the gamete known as sperm, which fuses with the larger female gamete, or ovum, in the process of fertilisation. A male organism cannot reproduce sexually without access to at least one ovum from a female, but some organisms can reproduce both sexually and asexually. Most male mammals, including male humans, have a Y chromosome, which codes for the production of larger amounts of testosterone to develop male reproductive organs.

XX male syndrome, also known as de la Chapelle syndrome, is a rare condition in which an individual with a 46,XX karyotype develops a male phenotype. Synonyms for XX male syndrome include 46,XX testicular difference of sex development
The ZW sex-determination system is a chromosomal system that determines the sex of offspring in birds, some fish and crustaceans such as the giant river prawn, some insects, the schistosome family of flatworms, and some reptiles, e.g. majority of snakes, lacertid lizards and monitors, including Komodo dragons. It is also present in some plants, where it has probably evolved independently on several occasions. The letters Z and W are used to distinguish this system from the XY sex-determination system. In the ZW system, females have a pair of dissimilar ZW chromosomes, and males have two similar ZZ chromosomes.
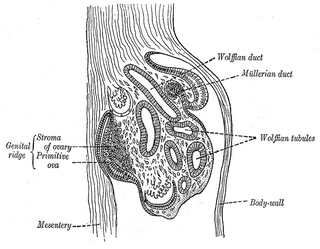
The genital ridge is the precursor to the gonads. The genital ridge initially consists mainly of mesenchyme and cells of underlying mesonephric origin. Once oogonia enter this area they attempt to associate with these somatic cells. Development proceeds and the oogonia become fully surrounded by a layer of cells.

Gonadal dysgenesis is classified as any congenital developmental disorder of the reproductive system in humans. It is atypical development of gonads in an embryo. One type of gonadal dysgenesis is the development of functionless, fibrous tissue, termed streak gonads, instead of reproductive tissue. Streak gonads are a form of aplasia, resulting in hormonal failure that manifests as sexual infantism and infertility, with no initiation of puberty and secondary sex characteristics.

Sexual differentiation in humans is the process of development of sex differences in humans. It is defined as the development of phenotypic structures consequent to the action of hormones produced following gonadal determination. Sexual differentiation includes development of different genitalia and the internal genital tracts and body hair plays a role in sex identification.

A hereditary carrier, is a person or other organism that has inherited a recessive allele for a genetic trait or mutation but usually does not display that trait or show symptoms of the disease. Carriers are, however, able to pass the allele onto their offspring, who may then express the genetic trait.
46,XX/46,XY is a chimeric genetic condition characterized by the presence of some cells that express a 46,XX karyotype and some cells that express a 46,XY karyotype in a single human being. The cause of the condition lies in utero with the aggregation of two distinct blastocysts or zygotes into a single embryo, which subsequently leads to the development of a single individual with two distinct cell lines, instead of a pair of fraternal twins. 46,XX/46,XY chimeras are the result of the merging of two non-identical twins. This is not to be confused with mosaicism or hybridism, neither of which are chimeric conditions.

Silene is a flowering plant genus that has evolved a dioecious reproductive system. This is made possible through heteromorphic sex chromosomes expressed as XY. Silene recently evolved sex chromosomes 5-10 million years ago and are widely used by geneticists and biologists to study the mechanisms of sex determination since they are one of only 39 species across 14 families of angiosperm that possess sex-determining genes. Silene are studied because of their ability to produce offspring with a plethora of reproductive systems. The common inference drawn from such studies is that the sex of the offspring is determined by the Y chromosome.


