Related Research Articles

The cerebellum is a major feature of the hindbrain of all vertebrates. Although usually smaller than the cerebrum, in some animals such as the mormyrid fishes it may be as large as or even larger. In humans, the cerebellum plays an important role in motor control. It may also be involved in some cognitive functions such as attention and language as well as emotional control such as regulating fear and pleasure responses, but its movement-related functions are the most solidly established. The human cerebellum does not initiate movement, but contributes to coordination, precision, and accurate timing: it receives input from sensory systems of the spinal cord and from other parts of the brain, and integrates these inputs to fine-tune motor activity. Cerebellar damage produces disorders in fine movement, equilibrium, posture, and motor learning in humans.
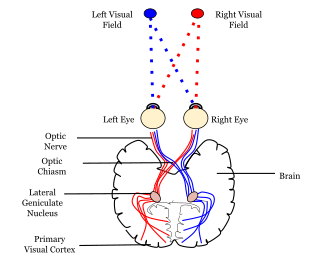
A neural pathway is the connection formed by axons that project from neurons to make synapses onto neurons in another location, to enable a signal to be sent from one region of the nervous system to another. Neurons are connected by a single axon, or by a bundle of axons known as a nerve tract, or fasciculus. Shorter neural pathways are found within grey matter in the brain, whereas longer projections, made up of myelinated axons, constitute white matter.

Pyramidal cells, or pyramidal neurons, are a type of multipolar neuron found in areas of the brain including the cerebral cortex, the hippocampus, and the amygdala. Pyramidal neurons are the primary excitation units of the mammalian prefrontal cortex and the corticospinal tract. Pyramidal neurons are also one of two cell types where the characteristic sign, Negri bodies, are found in post-mortem rabies infection. Pyramidal neurons were first discovered and studied by Santiago Ramón y Cajal. Since then, studies on pyramidal neurons have focused on topics ranging from neuroplasticity to cognition.

The auditory system is the sensory system for the sense of hearing. It includes both the sensory organs and the auditory parts of the sensory system.

The lateral lemniscus is a tract of axons in the brainstem that carries information about sound from the cochlear nucleus to various brainstem nuclei and ultimately the contralateral inferior colliculus of the midbrain. Three distinct, primarily inhibitory, cellular groups are located interspersed within these fibers, and are thus named the nuclei of the lateral lemniscus.

Basket cells are inhibitory GABAergic interneurons of the brain, found throughout different regions of the cortex and cerebellum.

Purkinje cells, or Purkinje neurons, are a class of GABAergic inhibitory neurons located in the cerebellum. They are named after their discoverer, Czech anatomist Jan Evangelista Purkyně, who characterized the cells in 1839.
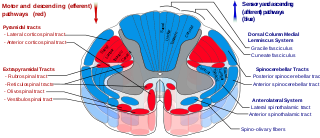
The spinocerebellar tract is a nerve tract originating in the spinal cord and terminating in the same side (ipsilateral) of the cerebellum.
An apical dendrite is a dendrite that emerges from the apex of a pyramidal cell. Apical dendrites are one of two primary categories of dendrites, and they distinguish the pyramidal cells from spiny stellate cells in the cortices. Pyramidal cells are found in the prefrontal cortex, the hippocampus, the entorhinal cortex, the olfactory cortex, and other areas. Dendrite arbors formed by apical dendrites are the means by which synaptic inputs into a cell are integrated. The apical dendrites in these regions contribute significantly to memory, learning, and sensory associations by modulating the excitatory and inhibitory signals received by the pyramidal cells.

The dentate nucleus is a cluster of neurons, or nerve cells, in the central nervous system that has a dentate – tooth-like or serrated – edge. It is located within the deep white matter of each cerebellar hemisphere, and it is the largest single structure linking the cerebellum to the rest of the brain. It is the largest and most lateral, or farthest from the midline, of the four pairs of deep cerebellar nuclei, the others being the globose and emboliform nuclei, which together are referred to as the interposed nucleus, and the fastigial nucleus. The dentate nucleus is responsible for the planning, initiation and control of voluntary movements. The dorsal region of the dentate nucleus contains output channels involved in motor function, which is the movement of skeletal muscle, while the ventral region contains output channels involved in nonmotor function, such as conscious thought and visuospatial function.
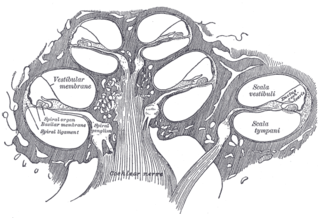
The cochlear nerve is one of two parts of the vestibulocochlear nerve, a cranial nerve present in amniotes, the other part being the vestibular nerve. The cochlear nerve carries auditory sensory information from the cochlea of the inner ear directly to the brain. The other portion of the vestibulocochlear nerve is the vestibular nerve, which carries spatial orientation information to the brain from the semicircular canals, also known as semicircular ducts.

The cochlear nuclear (CN) complex comprises two cranial nerve nuclei in the human brainstem, the ventral cochlear nucleus (VCN) and the dorsal cochlear nucleus (DCN). The ventral cochlear nucleus is unlayered whereas the dorsal cochlear nucleus is layered. Auditory nerve fibers, fibers that travel through the auditory nerve carry information from the inner ear, the cochlea, on the same side of the head, to the nerve root in the ventral cochlear nucleus. At the nerve root the fibers branch to innervate the ventral cochlear nucleus and the deep layer of the dorsal cochlear nucleus. All acoustic information thus enters the brain through the cochlear nuclei, where the processing of acoustic information begins. The outputs from the cochlear nuclei are received in higher regions of the auditory brainstem.

The dorsal cochlear nucleus, is a cortex-like structure on the dorso-lateral surface of the brainstem. Along with the ventral cochlear nucleus (VCN), it forms the cochlear nucleus (CN), where all auditory nerve fibers from the cochlea form their first synapses.

Hippocampus anatomy describes the physical aspects and properties of the hippocampus, a neural structure in the medial temporal lobe of the brain. It has a distinctive, curved shape that has been likened to the sea-horse monster of Greek mythology and the ram's horns of Amun in Egyptian mythology. This general layout holds across the full range of mammalian species, from hedgehog to human, although the details vary. For example, in the rat, the two hippocampi look similar to a pair of bananas, joined at the stems. In primate brains, including humans, the portion of the hippocampus near the base of the temporal lobe is much broader than the part at the top. Due to the three-dimensional curvature of this structure, two-dimensional sections such as shown are commonly seen. Neuroimaging pictures can show a number of different shapes, depending on the angle and location of the cut.

In neurophysiology, a dendritic spike refers to an action potential generated in the dendrite of a neuron. Dendrites are branched extensions of a neuron. They receive electrical signals emitted from projecting neurons and transfer these signals to the cell body, or soma. Dendritic signaling has traditionally been viewed as a passive mode of electrical signaling. Unlike its axon counterpart which can generate signals through action potentials, dendrites were believed to only have the ability to propagate electrical signals by physical means: changes in conductance, length, cross sectional area, etc. However, the existence of dendritic spikes was proposed and demonstrated by W. Alden Spencer, Eric Kandel, Rodolfo Llinás and coworkers in the 1960s and a large body of evidence now makes it clear that dendrites are active neuronal structures. Dendrites contain voltage-gated ion channels giving them the ability to generate action potentials. Dendritic spikes have been recorded in numerous types of neurons in the brain and are thought to have great implications in neuronal communication, memory, and learning. They are one of the major factors in long-term potentiation.
The anatomy of the cerebellum can be viewed at three levels. At the level of gross anatomy, the cerebellum consists of a tightly folded and crumpled layer of cortex, with white matter underneath, several deep nuclei embedded in the white matter, and a fluid-filled ventricle in the middle. At the intermediate level, the cerebellum and its auxiliary structures can be broken down into several hundred or thousand independently functioning modules or compartments known as microzones. At the microscopic level, each module consists of the same small set of neuronal elements, laid out with a highly stereotyped geometry.
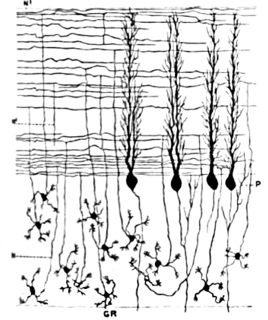
Cerebellar granule cells form the thick granular layer of the cerebellar cortex and are among the smallest neurons in the brain. Cerebellar granule cells are also the most numerous neurons in the brain: in humans, estimates of their total number average around 50 billion, which means that they constitute about 3/4 of the brain's neurons.
The name granule cell has been used for a number of different types of neurons whose only common feature is that they all have very small cell bodies. Granule cells are found within the granular layer of the cerebellum, the dentate gyrus of the hippocampus, the superficial layer of the dorsal cochlear nucleus, the olfactory bulb, and the cerebral cortex.
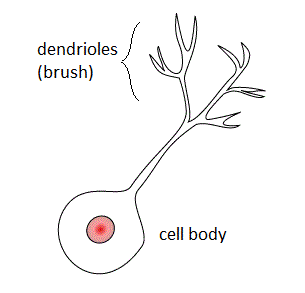
Unipolar brush cells (UBCs) are a class of excitatory glutamatergic interneuron found in the granular layer of the cerebellar cortex and also in the granule cell domain of the cochlear nucleus.
Lugaro cells are primary sensory interneurons of the cerebellum, that have an inhibitory function. They are fusiform, having a spindle shape that tapers at each end. They were first described by Ernesto Lugaro in the early 20th century. Lugaro cells are found just beneath the layer of Purkinje cells between the molecular layer and the granular layer. They have thick principal dendrites coming from opposite poles of their bodies. These dendrites are very long and travel along the boundary between the Purkinje layer and the granular layer. They seem to contact from 5 to 15 Purkinje cells in a horizontal direction. They can also sense stimuli close to the Purkinje cells and their dendrites form a large receptive area that monitors the environment near to the Purkinje cells. Whilst their dendrites make contact with the Purkinje cells they also receive inputs from branches of the Purkinje axons by which they seem to have a sampling and integration role.
References
- ↑ Mugnaini, E; Morgan, JI (Dec 1987). "The neuropeptide cerebellin is a marker for two similar neuronal circuits in rat brain". Proceedings of the National Academy of Sciences of the United States of America. 84 (23): 8692–6. Bibcode:1987PNAS...84.8692M. doi: 10.1073/pnas.84.23.8692 . PMC 299612 . PMID 3317418.
- ↑ Mugnaini, E; Berrebi, AS; Dahl, AL; Morgan, JI (Dec 1987). "The polypeptide PEP-19 is a marker for Purkinje neurons in cerebellar cortex and cartwheel neurons in the dorsal cochlear nucleus". Archives Italiennes de Biologie. 126 (1): 41–67. PMID 3449006.
- ↑ "Home". neurolex.org.
- ↑ Berrebi, AS; Mugnaini, E (1991). "Distribution and targets of the cartwheel cell axon in the dorsal cochlear nucleus of the guinea pig". Anatomy and Embryology. 183 (5): 427–54. doi:10.1007/BF00186433. PMID 1862946. S2CID 22783529.