Related Research Articles

The Ichneumonidae, also known as the ichneumon wasps, Darwin wasps, or ichneumonids, are a family of parasitoid wasps of the insect order Hymenoptera. They are one of the most diverse groups within the Hymenoptera with roughly 25,000 species currently described. However, this likely represents less than a quarter of their true richness as reliable estimates are lacking, along with much of the most basic knowledge about their ecology, distribution, and evolution. Ichneumonid wasps, with very few exceptions, attack the immature stages of holometabolous insects and spiders, eventually killing their hosts. They thus fulfill an important role as regulators of insect populations, both in natural and semi-natural systems, making them promising agents for biological control.

Bergmann's rule is an ecogeographical rule that states that within a broadly distributed taxonomic clade, populations and species of larger size are found in colder environments, while populations and species of smaller size are found in warmer regions. Bergmann's rule only describes the overall size of the animals, but does not include body parts like Allen's rule does.
The metabolic theory of ecology (MTE) is the ecological component of the more general Metabolic Scaling Theory and Kleiber's law. It posits that the metabolic rate of organisms is the fundamental biological rate that governs most observed patterns in ecology. MTE is part of a larger set of theory known as metabolic scaling theory that attempts to provide a unified theory for the importance of metabolism in driving pattern and process in biology from the level of cells all the way to the biosphere.
Macroecology is the subfield of ecology that deals with the study of relationships between organisms and their environment at large spatial scales to characterise and explain statistical patterns of abundance, distribution and diversity. The term was coined in a small monograph published in Spanish in 1971 by Guillermo Sarmiento and Maximina Monasterio, two Venezuelan researchers working in tropical savanna ecosystems and later used by James Brown of the University of New Mexico and Brian Maurer of Michigan State University in a 1989 paper in Science.
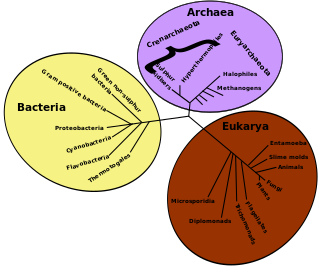
Evolutionary ecology lies at the intersection of ecology and evolutionary biology. It approaches the study of ecology in a way that explicitly considers the evolutionary histories of species and the interactions between them. Conversely, it can be seen as an approach to the study of evolution that incorporates an understanding of the interactions between the species under consideration. The main subfields of evolutionary ecology are life history evolution, sociobiology, the evolution of interspecific interactions and the evolution of biodiversity and of ecological communities.
Rapoport's rule is an ecogeographical rule that states that latitudinal ranges of plants and animals are generally smaller at lower latitudes than at higher latitudes.

The intermediate disturbance hypothesis (IDH) suggests that local species diversity is maximized when ecological disturbance is neither too rare nor too frequent. At low levels of disturbance, more competitive organisms will push subordinate species to extinction and dominate the ecosystem. At high levels of disturbance, due to frequent forest fires or human impacts like deforestation, all species are at risk of going extinct. According to IDH theory, at intermediate levels of disturbance, diversity is thus maximized because species that thrive at both early and late successional stages can coexist. IDH is a nonequilibrium model used to describe the relationship between disturbance and species diversity. IDH is based on the following premises: First, ecological disturbances have major effects on species richness within the area of disturbance. Second, interspecific competition results in one species driving a competitor to extinction and becoming dominant in the ecosystem. Third, moderate ecological scale disturbances prevent interspecific competition.

Species richness, or biodiversity, increases from the poles to the tropics for a wide variety of terrestrial and marine organisms, often referred to as the latitudinal diversity gradient (LDG). The LDG is one of the most widely recognized patterns in ecology. The LDG has been observed to varying degrees in Earth's past. A parallel trend has been found with elevation, though this is less well-studied.
Thorson's rule is an ecogeographical rule which states that benthic marine invertebrates at low latitudes tend to produce large numbers of eggs developing to pelagic and widely dispersing larvae, whereas at high latitudes such organisms tend to produce fewer and larger lecithotrophic (yolk-feeding) eggs and larger offspring, often by viviparity or ovoviviparity, which are often brooded.
The issue of what exactly defines a vacant niche, also known as empty niche, and whether they exist in ecosystems is controversial. The subject is intimately tied into a much broader debate on whether ecosystems can reach equilibrium, where they could theoretically become maximally saturated with species. Given that saturation is a measure of the number of species per resource axis per ecosystem, the question becomes: is it useful to define unused resource clusters as niche 'vacancies'?

The body size-species richness distribution is a pattern observed in the way taxa are distributed over large spatial scales. The number of species that exhibit small body size generally far exceed the number of species that are large-bodied. Macroecology has long sought to understand the mechanisms that underlie the patterns of biodiversity, such as the body size-species richness pattern.

Klaus Rohde is a German biologist at the University of New England (UNE), Australia, known particularly for his work on marine parasitology, evolutionary ecology/zoogeography, and phylogeny/ultrastructure of lower invertebrates.
The Janzen–Connell hypothesis is a widely accepted explanation for the maintenance of tree species biodiversity in tropical rainforests. It was published independently in the early 1970s by Daniel Janzen and Joseph Connell. According to their hypothesis, host-specific herbivores, pathogens, or other natural enemies make the areas near a parent tree inhospitable for the survival of seedlings. These natural enemies are referred to as 'distance-responsive predators' if they kill seeds or seedlings near the parent tree, or 'density-dependent predators' if they kill seeds or seedlings where they are most abundant. Such predators can prevent any one species from dominating the landscape, because if that species is too common, there will be few safe places for its seedlings to survive. However, because the predators are host-specific, they will not harm other tree species. As a result, if a species becomes very rare, then more predator-free areas will become available, giving that species' seedlings a competitive advantage. This negative feedback allows the tree species to coexist, and can be classified as a stabilizing mechanism.

This is a list of topics in biodiversity.

Elevational diversity gradient (EDG) is an ecological pattern where biodiversity changes with elevation. The EDG states that species richness tends to increase as elevation increases, up to a certain point, creating a "diversity bulge" at middle elevations. There have been multiple hypotheses proposed for explaining the EDG, none of which accurately describe the phenomenon in full.

Clutch size refers to the number of eggs laid in a single brood by a nesting pair of birds. The numbers laid by a particular species in a given location are usually well defined by evolutionary trade-offs with many factors involved, including resource availability and energetic constraints. Several patterns of variation have been noted and the relationship between latitude and clutch size has been a topic of interest in avian reproduction and evolution. David Lack and R.E. Moreau were among the first to investigate the effect of latitude on the number of eggs per nest. Since Lack's first paper in the mid-1940s there has been extensive research on the pattern of increasing clutch size with increasing latitude. The proximate and ultimate causes for this pattern have been a subject of intense debate involving the development of ideas on group, individual, and gene-centric views of selection.
The term phylogenetic niche conservatism has seen increasing use in recent years in the scientific literature, though the exact definition has been a matter of some contention. Fundamentally, phylogenetic niche conservatism refers to the tendency of species to retain their ancestral traits. When defined as such, phylogenetic niche conservatism is therefore nearly synonymous with phylogenetic signal. The point of contention is whether or not "conservatism" refers simply to the tendency of species to resemble their ancestors, or implies that "closely related species are more similar than expected based on phylogenetic relationships". If the latter interpretation is employed, then phylogenetic niche conservatism can be seen as an extreme case of phylogenetic signal, and implies that the processes which prevent divergence are in operation in the lineage under consideration. Despite efforts by Jonathan Losos to end this habit, however, the former interpretation appears to frequently motivate scientific research. In this case, phylogenetic niche conservatism might best be considered a form of phylogenetic signal reserved for traits with broad-scale ecological ramifications. Thus, phylogenetic niche conservatism is usually invoked with regards to closely related species occurring in similar environments.

Fecundity selection, also known as fertility selection, is the fitness advantage resulting from the preference of traits that increase the number of offspring. Charles Darwin formulated the theory of fecundity selection between 1871 and 1874 to explain the widespread evolution of female-biased sexual size dimorphism (SSD), where females were larger than males.
Countergradient variation is a type of phenotypic plasticity that occurs when the phenotypic variation determined by a biological population's genetic components opposes the phenotypic variation caused by an environmental gradient. This can cause different populations of the same organism to display similar phenotypes regardless of their underlying genetics and differences in their environments.

Roberto Cazzolla Gatti is an Italian ecologist and evolutionary biologist with a Ph.D. in forest ecology. His research deals with Biological Diversity and Conservation, and Global Environmental Protection occurring, therefore, at the interface between macroecology, evolutionary biology, and behavioural ecology.
References
- 1 2 K. Rohde: Latitudinal gradients in species diversity: the search for the primary cause, Oikos, 65, 514-527,1992.
- ↑ K. Rohde: Latitudinal gradients in species diversity and their causes. I. A review of the hypotheses explaining the gradients. Biologisches Zentralblatt 97, 393-403, 1978a.
- ↑ K. Rohde: Latitudinal gradients in species diversity and their causes. II. Marine parasitological evidence for a time hypothesis. Biologisches Zentralblatt 97, 405-418, 1978b.
- ↑ B. Rensch: Neuere Probleme der Abstammungslehre. Die transspezifische Evolution. Encke, Stuttgart, 1954.
- ↑ R.E. Ricklefs: Ecology. Nelson and Sons, London, 1973.
- ↑ F.G. Stehli, E.G. Douglas and N.D. Newell: Generation and maintenance of gradients in taxonomic diversity. Science 164, 947-949, 1969.
- ↑ N.W. Timofeeff-Ressovsky, K.G. Zimmer und M. Delbrück: Über die Natur der Genmutation und der Genstruktur. Nachrichten aus der Biologie der Gesellschaft der Wissenschaften Göttingen I, 189-245, 1935.
- 1 2 K. Rohde: Nonequilibrium Ecology , Cambridge University Press, Cambridge, 2005b, 223 pp. ISBN 0-521-67455-7.
- ↑ D.Jablonski: The future of the fossil record, Science 284, 2114-2116, 1999.
- ↑ M. Cardillo: Latitude and rates of diversification in birds and butterflies. Proceedings of the Royal Society London 266, 1221-1225,1999.
- ↑ A.P. Allen, J.H. Brown, and J.F. Gillooly: Global biodiversity, biochemical kinetics, and the energetic-equivalence rule. Science, 297, 1545-1548, 2002.
- ↑ J.F. Gillooly, E.L. Charnov, G.B. West, M.Van Savage, and J.H. Brown: Effects of size and temperature on developmental time. Nature 417, 70–73, 2002.
- ↑ J.H. Brown, J.F. Gillooly, A.P. Allen, M. Van Savage, and G.. West,. (2004). Toward a metabolic theory of ecology. Ecology 85, 1771-1789.
- ↑ C. Bazin, P. Capy, D. Higuet, and T. Langin, T.: Séquences d’AND mobiles et évolution du génome. Pour Sci., Hors. Sér. Janvier 97, 106-109., 1997 (zitiert in. Harmelin-Vivien 2002).
- ↑ M.L. Harmelin-Vivien: Energetics and fish diversity on coral reefs. In: Sale, P.F. Hrsg. Coral reef fishes. Dynamics and diversity in a complex ecosystem. Academic Press, Amsterdam, pp. 265-274, 2002.
- ↑ M. Kaspari, P.S. Ward and M.Yuan: Energy gradients and the geographical distribution of local ant diversity. Oecologia 140, 407-413, 2004.
- ↑ K. Rohde and D. Stauffer: "Simulation of geographical trends in Chowdhury ecosystem model", Advances in Complex Systems 8, 451-464, 2005.
- ↑ K. Rohde: Eine neue Ökologie. Aktuelle Probleme der evolutionären Ökologie". Naturwissenschaftliche Rundschau, 58, 420-426, 2005.
- ↑ S. Wright, J. Keeling and L. Gillman 2006. The road from Santa Rosalia: a faster tempo of evolution in tropical climates. Proceedings of the National Academy of Sciences 103, 7718 –7722.
- ↑ Gillman, L.N., Keeling, D.J., Ross,H.A. and Wright, S.D. 2009. Latitude, elevation and the tempo of molecular evolution in mammals. Proceedings of the Royal Society B 2009.
- ↑ S. Wright, J. Keeling and L. Gillman. 2006. The road from Santa Rosalia: a faster tempo of evolution in tropical climates. Proceedings of the National Academy of Sciences 103, 7718 –7722.
- ↑ Gillman L.N., Keeling D.J., Gardner R.C., Wright S.D. Faster evolution of highly conserved DNA in tropical plants. Journal of evolutionary Biology 23, 1327-1330,2010
- ↑ Wright S.D., Gillman L.N., Ross H.A. and Keeling D.J. Energy and tempo of evolution in amphibians. Global Ecology and Biogeography 2010
- ↑ Vázquez, D.P. and Stevens, R.D. The latitudinal gradient in niche breadth: concepts and evidence. American Naturalist 164, E1- E19, 2004.
- ↑ J. C.Briggs. Species diversity: land and sea compared. Systematic Biology 43, 130-135, 1994.
- ↑ J F. Grassle and N .J. Maciolek: Deepsea species richness: regional and local diversity estimates from, quantitative bottom samples. American Naturalist 139, 313-341, 1992.
- ↑ K. Rohde: Ecology and biogeography of marine parasites. Advances in marine biology 43,1-86, 2002.