Related Research Articles

The cerebral cortex, also known as the cerebral mantle, is the outer layer of neural tissue of the cerebrum of the brain in humans and other mammals. The cerebral cortex mostly consists of the six-layered neocortex, with just 10% consisting of the allocortex. It is separated into two cortices, by the longitudinal fissure that divides the cerebrum into the left and right cerebral hemispheres. The two hemispheres are joined beneath the cortex by the corpus callosum. The cerebral cortex is the largest site of neural integration in the central nervous system. It plays a key role in attention, perception, awareness, thought, memory, language, and consciousness. The cerebral cortex is part of the brain responsible for cognition.

Brodmann area 23 (BA23) is a region in the brain that lies inside the posterior cingulate cortex. It lies between Brodmann area 30 and Brodmann area 31 and is located on the medial wall of the cingulate gyrus between the callosal sulcus and the cingulate sulcus.

Brodmann area 8 is one of Brodmann's cytologically defined regions of the brain. It is involved in planning complex movements.

Brodmann area 10 is the anterior-most portion of the prefrontal cortex in the human brain. BA10 was originally defined broadly in terms of its cytoarchitectonic traits as they were observed in the brains of cadavers, but because modern functional imaging cannot precisely identify these boundaries, the terms anterior prefrontal cortex, rostral prefrontal cortex and frontopolar prefrontal cortex are used to refer to the area in the most anterior part of the frontal cortex that approximately covers BA10—simply to emphasize the fact that BA10 does not include all parts of the prefrontal cortex.

Brodmann area 5 is one of Brodmann's cytoarchitectural defined regions of the brain. It is involved in somatosensory processing, movement and association, and is part of the posterior parietal cortex.

Brodmann area 19, or BA 19, is part of the occipital lobe cortex in the human brain. Along with area 18, it comprises the extrastriate cortex. In humans with normal sight, extrastriate cortex is a visual association area, with feature-extracting, shape recognition, attentional, and multimodal integrating functions.

Brodmann area 20, or BA20, is part of the temporal cortex in the human brain. The region encompasses most of the ventral temporal cortex, a region believed to play a part in high-level visual processing and recognition memory.

Brodmann area 21, or BA21, is part of the temporal cortex in the human brain. The region encompasses most of the lateral temporal cortex and is also known as middle temporal area 21. In the human it corresponds approximately to the middle temporal gyrus.

Brodmann area 11 is one of Brodmann's cytologically defined regions of the brain. It is in the orbitofrontal cortex which is above the eye sockets (orbitae). It is involved in decision making, processing rewards, and encoding new information into long-term memory.

Brodmann area 4 refers to the primary motor cortex of the human brain. It is located in the posterior portion of the frontal lobe.

Brodmann area 24 is part of the anterior cingulate in the human brain.

The Brodmann area 32, also known in the human brain as the dorsal anterior cingulate area 32, refers to a subdivision of the cytoarchitecturally defined cingulate cortex. In the human it forms an outer arc around the anterior cingulate gyrus. The cingulate sulcus defines approximately its inner boundary and the superior rostral sulcus (H) its ventral boundary; rostrally it extends almost to the margin of the frontal lobe. Cytoarchitecturally it is bounded internally by the ventral anterior cingulate area 24, externally by medial margins of the agranular frontal area 6, intermediate frontal area 8, granular frontal area 9, frontopolar area 10, and prefrontal area 11-1909. (Brodmann19-09).
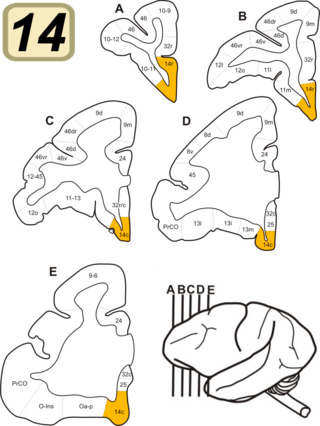
Brodmann Area 14 is one of Brodmann's subdivisions of the cerebral cortex in the brain. It was defined by Brodmann in the guenon monkey . While Brodmann, writing in 1909, argued that no equivalent structure existed in humans, later work demonstrated that area 14 has a clear homologue in the human ventromedial prefrontal cortex.

Brodmann area 12 is a subdivision of the cerebral cortex of the guenon defined on the basis of cytoarchitecture. It occupies the most rostral portion of the frontal lobe. Brodmann-1909 did not regard it as homologous, either topographically or cytoarchitecturally, to rostral area 12 of the human. Distinctive features (Brodmann-1905): a quite distinct internal granular layer (IV) separates slender pyramidal cells of the external pyramidal layer (III) and the internal pyramidal layer (V); the multiform layer (VI) is expanded, contains widely dispersed spindle cells and merges gradually with the underlying cortical white matter; all cells, including the pyramidal cells of the external and internal pyramidal layers are inordinately small; the internal pyramidal layer (V) also contains spindle cells in groups of two to five located close to its border with the internal granular layer (IV).
Brodmann Area 15 is one of Brodmann's subdivisions of the cerebral cortex in the brain.
Brodmann area 16 is a subdivision of the cerebral cortex of the guenon defined on the basis of cytoarchitecture. It is a relatively undifferentiated cortical area that Brodmann regarded as part of the insula because of the relation of its innermost multiform layer (VI) with the claustrum (VICl). The laminar organization of cortex is almost totally lacking. The molecular layer (I) is wide as in area 15 of Brodmann-1905. The space between layer I and layer VI is composed of a mixture of pyramidal cells and spindle cells with no significant number of granule cells. Pyramidal cells clump in the outer part to form glomeruli similar to those seen in some of the primary olfactory areas (Brodmann-1905).
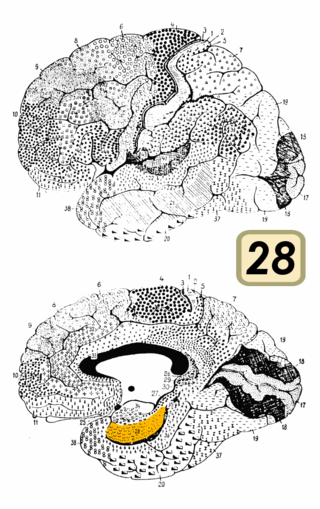
Brodmann area 28 is a subdivision of the cerebral cortex defined on the basis of cytoarchitecture. It is located on the medial aspect of the temporal lobe and is part of the entorhinal cortex (Brodmann-1909).
The trisynaptic circuit, or trisynaptic loop is a relay of synaptic transmission in the hippocampus. The circuit was initially described by the neuroanatomist Santiago Ramon y Cajal, in the early twentieth century, using the Golgi staining method. After the discovery of the trisynaptic circuit, a series of research has been conducted to determine the mechanisms driving this circuit. Today, research is focused on how this loop interacts with other parts of the brain, and how it influences human physiology and behaviour. For example, it has been shown that disruptions within the trisynaptic circuit lead to behavioural changes in rodent and feline models.
Heterotypic cortex consists of those areas of the mature neocortex that deviate markedly from the homogeneous six-layered internal structure seen in the third trimester of human gestation. A few neocortical areas, such as Brodmann area 17 and the granular insular cortex, undergo modification to more than six layers; and in a few areas, such as Brodmann area 4, the number of layers is reduced. Heterotypic cortex is contrasted to homotypic cortex, which retains the fetal six-layered pattern into adulthood. The number of heterotypic areas is small and the specific areas differ somewhat by species.
References
- ↑ Brodmann, K (2005). Localisation in the cerebral cortex. Imperial College Press. pp. 50–52. ISBN 0387269177.