
Drosophila is a genus of flies, belonging to the family Drosophilidae, whose members are often called "small fruit flies" or pomace flies, vinegar flies, or wine flies, a reference to the characteristic of many species to linger around overripe or rotting fruit. They should not be confused with the Tephritidae, a related family, which are also called fruit flies ; tephritids feed primarily on unripe or ripe fruit, with many species being regarded as destructive agricultural pests, especially the Mediterranean fruit fly.

Drosophila melanogaster is a species of fly in the family Drosophilidae. The species is often referred to as the fruit fly or lesser fruit fly, or less commonly the "vinegar fly", "pomace fly", or "banana fly". In the wild, D. melanogaster are attracted to rotting fruit and fermenting beverages, and are often found in orchards, kitchens and pubs.

Sperm competition is the competitive process between spermatozoa of two or more different males to fertilize the same egg during sexual reproduction. Competition can occur when females have multiple potential mating partners. Greater choice and variety of mates increases a female's chance to produce more viable offspring. However, multiple mates for a female means each individual male has decreased chances of producing offspring. Sperm competition is an evolutionary pressure on males, and has led to the development of adaptations to increase male's chance of reproductive success. Sperm competition results in a sexual conflict between males and females. Males have evolved several defensive tactics including: mate-guarding, mating plugs, and releasing toxic seminal substances to reduce female re-mating tendencies to cope with sperm competition. Offensive tactics of sperm competition involve direct interference by one male on the reproductive success of another male, for instance by mate guarding or by physically removing another male's sperm prior to mating with a female. For an example, see Gryllus bimaculatus.
Interlocus contest evolution (ICE) is a process of intergenomic conflict by which different loci within a single genome antagonistically coevolve. ICE supposes that the Red Queen process, which is characterized by a never-ending antagonistic evolutionary arms race, does not only apply to species but also to genes within the genome of a species.
The mechanisms of reproductive isolation are a collection of evolutionary mechanisms, behaviors and physiological processes critical for speciation. They prevent members of different species from producing offspring, or ensure that any offspring are sterile. These barriers maintain the integrity of a species by reducing gene flow between related species.

Sexual conflict or sexual antagonism occurs when the two sexes have conflicting optimal fitness strategies concerning reproduction, particularly over the mode and frequency of mating, potentially leading to an evolutionary arms race between males and females. In one example, males may benefit from multiple matings, while multiple matings may harm or endanger females, due to the anatomical differences of that species. Sexual conflict underlies the evolutionary distinction between male and female.

The sexy son hypothesis in evolutionary biology and sexual selection, proposed by Patrick J. Weatherhead and Raleigh J. Robertson of Queen's University in Kingston, Ontario in 1979, states that a female's ideal mate choice among potential mates is one whose genes will produce males with the best chance of reproductive success. This implies that other benefits the father can offer the mother or offspring are less relevant than they may appear, including his capacity as a parental caregiver, territory and any nuptial gifts. Fisher's principle means that the sex ratio is always near 1:1 between males and females, yet what matters most are her "sexy sons'" future breeding successes, more likely if they have a promiscuous father, in creating large numbers of offspring carrying copies of her genes. This sexual selection hypothesis has been researched in species such as the European pied flycatcher.

Drosophila pseudoobscura is a species of fruit fly, used extensively in lab studies of speciation. It is native to western North America.

Scathophaga stercoraria, commonly known as the yellow dung fly or the golden dung fly, is one of the most familiar and abundant flies in many parts of the Northern Hemisphere. As its common name suggests, it is often found on the feces of large mammals, such as horses, cattle, sheep, deer, and wild boar, where it goes to breed. The distribution of S. stercoraria is likely influenced by human agriculture, especially in northern Europe and North America. The Scathophaga are integral in the animal kingdom due to their role in the natural decomposition of dung in fields. They are also very important in the scientific world due to their short life cycles and susceptibility to experimental manipulations; thus, they have contributed significant knowledge about animal behavior.
Bateman's principle, in evolutionary biology, is that in most species, variability in reproductive success is greater in males than in females. It was first proposed by Angus John Bateman (1919–1996), an English geneticist. Bateman suggested that, since males are capable of producing millions of sperm cells with little effort, while females invest much higher levels of energy in order to nurture a relatively small number of eggs, the female plays a significantly larger role in their offspring's reproductive success. Bateman's paradigm thus views females as the limiting factor of parental investment, over which males will compete in order to copulate successfully.
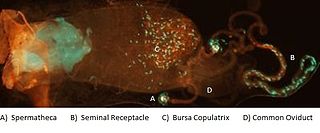
Female sperm storage is a biological process and often a type of sexual selection in which sperm cells transferred to a female during mating are temporarily retained within a specific part of the reproductive tract before the oocyte, or egg, is fertilized. This process takes place in some species of animals, but not in humans. The site of storage is variable among different animal taxa and ranges from structures that appear to function solely for sperm retention, such as insect spermatheca and bird sperm storage tubules, to more general regions of the reproductive tract enriched with receptors to which sperm associate before fertilization, such as the caudal portion of the cow oviduct containing sperm-associating annexins. Female sperm storage is an integral stage in the reproductive process for many animals with internal fertilization. It has several documented biological functions including:
Sexual antagonistic co-evolution is the relationship between males and females where sexual morphology changes over time to counteract the opposite's sex traits to achieve the maximum reproductive success. This has been compared to an arms race between sexes. In many cases, male mating behavior is detrimental to the female's fitness. For example, when insects reproduce by means of traumatic insemination, it is very disadvantageous to the female's health. During mating, males will try to inseminate as many females as possible, however, the more times a female's abdomen is punctured, the less likely she is to survive. Females that possess traits to avoid multiple matings will be more likely to survive, resulting in a change in morphology. In males, genitalia is relatively simple and more likely to vary among generations compared to female genitalia. This results in a new trait that females have to avoid in order to survive.
Intralocus sexual conflict is a type of sexual conflict that occurs when a genetic locus harbours alleles which have opposing effects on the fitness of each sex, such that one allele improves the fitness of males, while the alternative allele improves the fitness of females. Such "sexually antagonistic" polymorphisms are ultimately generated by two forces: (i) the divergent reproductive roles of each sex, such as conflicts over optimal mating strategy, and (ii) the shared genome of both sexes, which generates positive between-sex genetic correlations for most traits. In the long term, intralocus sexual conflict is resolved when genetic mechanisms evolve that decouple the between-sex genetic correlations between traits. This can be achieved, for example, via the evolution of sex-biased or sex-limited genes.
A behaviour mutation is a genetic mutation that alters genes that control the way in which an organism behaves, causing their behavioural patterns to change.
Cryptic female choice is a form of mate choice which occurs both in pre and post copulatory circumstances when females in certain species use physical or chemical mechanisms to control a male's success of fertilizing their ova or ovum; i.e. by selecting whether sperm are successful in fertilizing their eggs or not. It occurs in internally-fertilizing species and involves differential use of sperm by females when sperm are available in the reproductive tract.

Prochyliza xanthostoma, the waltzing fly, is a species of carrion-feeding cheese skipper, insects in the family Piophilidae and the order Diptera. P. xanthostoma is a member of the genus Prochyliza, which contains eleven species. The adult flies are found through North America and are brown-bodied, with orange and black coloring. Mating occurs on animal carcasses and male perform mating rituals; females engage in ejaculate feeding. The waltzing fly is known for its exaggerated sexual dimorphism and has thus become a prominent model for sexual dimorphism and larval behavior. These organisms are known as cheese skippers because when startled, the larvae can leap several inches into the air. P. xanthostoma is an important model organism for sexual selection, larval behavior, and adult reproductive success and survivability.

Seminal fluid proteins (SFPs) or accessory gland proteins (Acps) are one of the non-sperm components of semen. In many animals with internal fertilization, males transfer a complex cocktail of proteins in their semen to females during copulation. These seminal fluid proteins often have diverse, potent effects on female post-mating phenotypes. SFPs are produced by the male accessory glands.
Mariana Federica Wolfner is the Goldwin Smith Professor of molecular biology and genetics at Cornell University. Her research investigates sexual conflict in the fruit fly Drosophila melanogaster. She was elected a member of the National Academy of Sciences (NAS) in 2019 in recognition of her distinguished and continuing achievements in original research.
Derocephalus angusticollis is a fly in the family Neriidae. They are typically found on the east coast of Australia near rotting vegetation. Aggregating on the rotting bark of trees such as Acacia longifolia and other trees in New South Wales and southern Queensland. D. angusticollis flies found in the wild have accelerated speeds of development and age of mortality when compared to those in captivity. Derocephalus One characteristic of the neriid fly is that it demonstrates sexual dimorphism. Males have a larger build as well as exaggerated physical characteristics such as wider heads and longer limbs. Certain phenotypic characteristics are dependent on the diet of the parents.
Genetic incompatibility describes the process by which mating yields offspring that are nonviable, prone to disease, or genetically defective in some way. In nature, animals can ill afford to devote costly resources for little or no reward, ergo, mating strategies have evolved to allow females to choose or otherwise determine mates which are more likely to result in viable offspring.

