Related Research Articles

Heredity, also called inheritance or biological inheritance, is the passing on of traits from parents to their offspring; either through asexual reproduction or sexual reproduction, the offspring cells or organisms acquire the genetic information of their parents. Through heredity, variations between individuals can accumulate and cause species to evolve by natural selection. The study of heredity in biology is genetics.
Genomic imprinting is an epigenetic phenomenon that causes genes to be expressed or not, depending on whether they are inherited from the female or male parent. Genes can also be partially imprinted. Partial imprinting occurs when alleles from both parents are differently expressed rather than complete expression and complete suppression of one parent's allele. Forms of genomic imprinting have been demonstrated in fungi, plants and animals. In 2014, there were about 150 imprinted genes known in mice and about half that in humans. As of 2019, 260 imprinted genes have been reported in mice and 228 in humans.
Selfish genetic elements are genetic segments that can enhance their own transmission at the expense of other genes in the genome, even if this has no positive or a net negative effect on organismal fitness. Genomes have traditionally been viewed as cohesive units, with genes acting together to improve the fitness of the organism.

A sex-determination system is a biological system that determines the development of sexual characteristics in an organism. Most organisms that create their offspring using sexual reproduction have two common sexes and a few less common intersex variations.

Haldane's rule is an observation about the early stage of speciation, formulated in 1922 by the British evolutionary biologist J. B. S. Haldane, that states that if — in a species hybrid — only one sex is inviable or sterile, that sex is more likely to be the heterogametic sex. The heterogametic sex is the one with two different sex chromosomes; in therian mammals, for example, this is the male.
A mating system is a way in which a group is structured in relation to sexual behaviour. The precise meaning depends upon the context. With respect to animals, the term describes which males and females mate under which circumstances. Recognised systems include monogamy, polygamy, and promiscuity, all of which lead to different mate choice outcomes and thus these systems affect how sexual selection works in the species which practice them. In plants, the term refers to the degree and circumstances of outcrossing. In human sociobiology, the terms have been extended to encompass the formation of relationships such as marriage.

Evolution of sexual reproduction describes how sexually reproducing animals, plants, fungi and protists could have evolved from a common ancestor that was a single-celled eukaryotic species. Sexual reproduction is widespread in eukaryotes, though a few eukaryotic species have secondarily lost the ability to reproduce sexually, such as Bdelloidea, and some plants and animals routinely reproduce asexually without entirely having lost sex. The evolution of sexual reproduction contains two related yet distinct themes: its origin and its maintenance. Bacteria and Archaea (prokaryotes) have processes that can transfer DNA from one cell to another, but it is unclear if these processes are evolutionarily related to sexual reproduction in Eukaryotes. In eukaryotes, true sexual reproduction by meiosis and cell fusion is thought to have arisen in the last eukaryotic common ancestor, possibly via several processes of varying success, and then to have persisted.

An inversion is a chromosome rearrangement in which a segment of a chromosome becomes inverted within its original position. An inversion occurs when a chromosome undergoes a two breaks within the chromosomal arm, and the segment between the two breaks inserts itself in the opposite direction in the same chromosome arm. The breakpoints of inversions often happen in regions of repetitive nucleotides, and the regions may be reused in other inversions. Chromosomal segments in inversions can be as small as 1 kilobases or as large as 100 megabases. The number of genes captured by an inversion can range from a handful of genes to hundreds of genes. Inversions can happen either through ectopic recombination between repetitive sequences, or through chromosomal breakage followed by non-homologous end joining.
Intragenomic conflict refers to the evolutionary phenomenon where genes have phenotypic effects that promote their own transmission in detriment of the transmission of other genes that reside in the same genome. The selfish gene theory postulates that natural selection will increase the frequency of those genes whose phenotypic effects cause their transmission to new organisms, and most genes achieve this by cooperating with other genes in the same genome to build an organism capable of reproducing and/or helping kin to reproduce. The assumption of the prevalence of intragenomic cooperation underlies the organism-centered concept of inclusive fitness. However, conflict among genes in the same genome may arise both in events related to reproduction and altruism.
Meiotic drive is a type of intragenomic conflict, whereby one or more loci within a genome will affect a manipulation of the meiotic process in such a way as to favor the transmission of one or more alleles over another, regardless of its phenotypic expression. More simply, meiotic drive is when one copy of a gene is passed on to offspring more than the expected 50% of the time. According to Buckler et al., "Meiotic drive is the subversion of meiosis so that particular genes are preferentially transmitted to the progeny. Meiotic drive generally causes the preferential segregation of small regions of the genome".
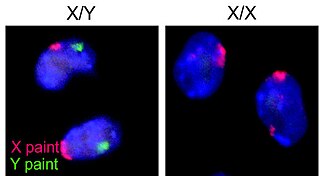
The ZW sex-determination system is a chromosomal system that determines the sex of offspring in birds, some fish and crustaceans such as the giant river prawn, some insects, the schistosome family of flatworms, and some reptiles, e.g. majority of snakes, lacertid lizards and monitors, including Komodo dragons. It is also present in some plants, where it has probably evolved independently on several occasions. The letters Z and W are used to distinguish this system from the XY sex-determination system. In the ZW system, females have a pair of dissimilar ZW chromosomes, and males have two similar ZZ chromosomes.

Haplodiploidy is a sex-determination system in which males develop from unfertilized eggs and are haploid, and females develop from fertilized eggs and are diploid. Haplodiploidy is sometimes called arrhenotoky.

Sexual conflict or sexual antagonism occurs when the two sexes have conflicting optimal fitness strategies concerning reproduction, particularly over the mode and frequency of mating, potentially leading to an evolutionary arms race between males and females. In one example, males may benefit from multiple matings, while multiple matings may harm or endanger females due to the anatomical differences of that species. Sexual conflict underlies the evolutionary distinction between male and female.
Mating types are the microorganism equivalent to sexes in multicellular lifeforms and are thought to be the ancestor to distinct sexes. They also occur in multicellular organisms such as fungi.
Sex-limited genes are genes that are present in both sexes of sexually reproducing species but are expressed in only one sex and have no penetrance, or are simply 'turned off' in the other. In other words, sex-limited genes cause the two sexes to show different traits or phenotypes, despite having the same genotype. This term is restricted to autosomal traits, and should not be confused with sex-linked characteristics, which have to do with genetic differences on the sex chromosomes. Sex-limited genes are also distinguished from sex-influenced genes, where the same gene will show differential expression in each sex. Sex-influenced genes commonly show a dominant/recessive relationship, where the same gene will have a dominant effect in one sex and a recessive effect in the other. However, the resulting phenotypes caused by sex-limited genes are present in only one sex and can be seen prominently in various species that typically show high sexual dimorphism.
Interlocus sexual conflict is a type of sexual conflict that occurs through the interaction of a set of antagonistic alleles at two or more different loci, or the location of a gene on a chromosome, in males and females, resulting in the deviation of either or both sexes from the fitness optima for the traits. A co-evolutionary arms race is established between the sexes in which either sex evolves a set of antagonistic adaptations that is detrimental to the fitness of the other sex. The potential for reproductive success in one organism is strengthened while the fitness of the opposite sex is weakened. Interlocus sexual conflict can arise due to aspects of male–female interactions such as mating frequency, fertilization, relative parental effort, female remating behavior, and female reproductive rate.
The theoretical foundations of evolutionary psychology are the general and specific scientific theories that explain the ultimate origins of psychological traits in terms of evolution. These theories originated with Charles Darwin's work, including his speculations about the evolutionary origins of social instincts in humans. Modern evolutionary psychology, however, is possible only because of advances in evolutionary theory in the 20th century.
Intralocus sexual conflict is a type of sexual conflict that occurs when a genetic locus harbours alleles which have opposing effects on the fitness of each sex, such that one allele improves the fitness of males, while the alternative allele improves the fitness of females. Such "sexually antagonistic" polymorphisms are ultimately generated by two forces: (i) the divergent reproductive roles of each sex, such as conflicts over optimal mating strategy, and (ii) the shared genome of both sexes, which generates positive between-sex genetic correlations for most traits. In the long term, intralocus sexual conflict is resolved when genetic mechanisms evolve that decouple the between-sex genetic correlations between traits. This can be achieved, for example, via the evolution of sex-biased or sex-limited genes.

Silene is a flowering plant genus that has evolved a dioecious reproductive system. This is made possible through heteromorphic sex chromosomes expressed as XY. Silene recently evolved sex chromosomes 5-10 million years ago and are widely used by geneticists and biologists to study the mechanisms of sex determination since they are one of only 39 species across 14 families of angiosperm that possess sex-determining genes. Silene are studied because of their ability to produce offspring with a plethora of reproductive systems. The common inference drawn from such studies is that the sex of the offspring is determined by the Y chromosome.
This glossary of genetics and evolutionary biology is a list of definitions of terms and concepts used in the study of genetics and evolutionary biology, as well as sub-disciplines and related fields, with an emphasis on classical genetics, quantitative genetics, population biology, phylogenetics, speciation, and systematics. It has been designed as a companion to Glossary of cellular and molecular biology, which contains many overlapping and related terms; other related glossaries include Glossary of biology and Glossary of ecology.
References
- 1 2 3 4 5 6 7 Rice & Holland, 1997. "The enemies within: intergenomic conflict, interlocus contest evolution (ICE), and the intraspecific Red Queen." Behavioral Ecology and Sociobiology 41(1): 1-10.
- ↑ Bateman, AJ. 1948. “Intra-sexual selection in Drosophila.” Heredity 2: 349-368.
- ↑ Trivers, R.L. 1972. "Parental investment and sexual selection Archived 2016-03-04 at the Wayback Machine ." Sexual Selection and the Descent of Man, 1871–1971. Campbell, Aldine Publishing. 136–179.
- ↑ Bonduriansky, R, SF Chenoweth. 2009. "Intralocus sexual conflict." Trends in Ecology & Evolution 24(5): 280-288.
- ↑ Ridley, M. 2000. Genome: The Autobiography of a Species in 23 Chapters. Harper and Collins.
- ↑ Bartholomew, GA, NE Collias. 1962. "The role of vocalization in the social behaviour of the northern elephant seal." Animal Behaviour 10(1): 7-14.