Related Research Articles

An annual plant is a plant that completes its life cycle, from germination to the production of seeds, within one growing season, and then dies. Globally, 6% of all plant species and 15% of herbaceous plants are annuals. The annual life cycle has independently emerged in over 120 different plant families throughout the entire angiosperm phylogeny.

A heterotroph is an organism that cannot produce its own food, instead taking nutrition from other sources of organic carbon, mainly plant or animal matter. In the food chain, heterotrophs are primary, secondary and tertiary consumers, but not producers. Living organisms that are heterotrophic include all animals and fungi, some bacteria and protists, and many parasitic plants. The term heterotroph arose in microbiology in 1946 as part of a classification of microorganisms based on their type of nutrition. The term is now used in many fields, such as ecology, in describing the food chain.

In ecology, the competitive exclusion principle, sometimes referred to as Gause's law, is a proposition that two species which compete for the same limited resource cannot coexist at constant population values. When one species has even the slightest advantage over another, the one with the advantage will dominate in the long term. This leads either to the extinction of the weaker competitor or to an evolutionary or behavioral shift toward a different ecological niche. The principle has been paraphrased in the maxim "complete competitors cannot coexist".
The metabolic theory of ecology (MTE) is the ecological component of the more general Metabolic Scaling Theory and Kleiber's law. It posits that the metabolic rate of organisms is the fundamental biological rate that governs most observed patterns in ecology. MTE is part of a larger set of theory known as metabolic scaling theory that attempts to provide a unified theory for the importance of metabolism in driving pattern and process in biology from the level of cells all the way to the biosphere.

Kleiber's law, named after Max Kleiber for his biology work in the early 1930s, states, after many observation that, for a vast number of animals, an animal's Basal Metabolic Rate scales to the 3⁄4 power of the animal's mass.

Allometry is the study of the relationship of body size to shape, anatomy, physiology and behaviour, first outlined by Otto Snell in 1892, by D'Arcy Thompson in 1917 in On Growth and Form and by Julian Huxley in 1932.

Robert Helmer MacArthur was a Canadian-born American ecologist who made a major impact on many areas of community and population ecology. He is considered to be one of the founders of ecology.
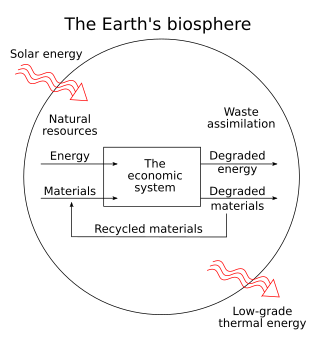
Metabolic rift is a theory of ecological crisis tendencies under the capitalist mode of production that sociologist John Bellamy Foster ascribes to Karl Marx. Quoting Marx, Foster defines this as the "irreparable rift in the interdependent process of social metabolism". Foster argues that Marx theorized a rupture in the metabolic interaction between humanity and the rest of nature emanating from capitalist agricultural production and the growing division between town and country.
The dynamic energy budget (DEB) theory is a formal metabolic theory which provides a single quantitative framework to dynamically describe the aspects of metabolism of all living organisms at the individual level, based on assumptions about energy uptake, storage, and utilization of various substances. The DEB theory adheres to stringent thermodynamic principles, is motivated by universally observed patterns, is non-species specific, and links different levels of biological organization as prescribed by the implications of energetics. Models based on the DEB theory have been successfully applied to over 1000 species with real-life applications ranging from conservation, aquaculture, general ecology, and ecotoxicology. The theory is contributing to the theoretical underpinning of the emerging field of metabolic ecology.
Dynamic reserve, in the context of the dynamic energy budget theory, refers to the set of metabolites that an organism can use for metabolic purposes. These chemical compounds can have active metabolic functions, however. They are not just "set apart for later use." Reserve differs from structure in the first place by its dynamics. Reserve has an implied turnover, because it is synthesized from food and used by metabolic processes occurring in cells. The turnover of structure depends on the maintenance of an organism. Maintenance is not required for reserve. A freshly laid egg consists almost exclusively of reserve, and hardly respires. The chemical compounds in the reserve have the same turnover, while that in the structure can have a different turnover, and so it depends on the compound.
A generalized compound is a mixture of chemical compounds of constant composition, despite possible changes in the total amount. The concept is used in the Dynamic Energy Budget theory, where biomass is partitioned into a limited set of generalised compounds, which contain a high percentage of organic compounds. The amount of generalized compound can be quantified in terms of weight, but more conveniently in terms of C-moles. The concept of strong homeostasis has an intimate relationship with that of generalised compound.
The DEBtox method for the evaluation of effects of toxicants makes use of the Dynamic Energy Budget (DEB) theory to quantify the effect. See the Organisation for Economic Co-operation and Development (OECD) report, below, for a description of the method.

Evolutionary physiology is the study of the biological evolution of physiological structures and processes; that is, the manner in which the functional characteristics of organisms have responded to natural selection or sexual selection or changed by random genetic drift across multiple generations during the history of a population or species. It is a sub-discipline of both physiology and evolutionary biology. Practitioners in the field come from a variety of backgrounds, including physiology, evolutionary biology, ecology, and genetics.
Jordan's rule is an ecogeographical rule that describes the inverse relationship between water temperature and meristic characteristics in various species of fish. The most commonly observed relationship is that fin ray, vertebrae, or scale numbers increase with decreasing temperature. The rule is named after David Starr Jordan (1851–1931), the father of American ichthyology.

Robert Eric Ricklefs is an American ornithologist and ecologist. He was the Curators' Professor of Biology at the University of Missouri, St. Louis from 1996 until August 2019.
In ecology, a priority effect refers to the impact that a particular species can have on community development as a result of its prior arrival at a site. There are two basic types of priority effects: inhibitory and facilitative. An inhibitory priority effect occurs when a species that arrives first at a site negatively affects a species that arrives later by reducing the availability of space or resources. In contrast, a facilitative priority effect occurs when a species that arrives first at a site alters abiotic or biotic conditions in ways that positively affect a species that arrives later. Inhibitory priority effects have been documented more frequently than facilitative priority effects. Studies indicate that both abiotic and biotic factors can affect the strength of priority effects.. Priority effects are a central and pervasive element of ecological community development that have significant implications for natural systems and ecological restoration efforts.

Capital breeding and income breeding refer to the methods by which some organisms perform time breeding and use resources to finance their breeding. The former "describes the situation in which reproduction is financed using stored capital; [whereas the latter] [...] refers to the use of concurrent intake to pay for a reproductive attempt."
Susan Patricia Harrison is a professor of ecology at the University of California, Davis who works on the dynamics of natural populations and ecological diversity. She is a fellow of the Ecological Society of America and the California Academy of Sciences. She has previously served as vice president of the American Society of Naturalists. She was elected to the National Academy of Sciences in 2018.
Taxon cycles refer to a biogeographical theory of how species evolve through range expansions and contractions over time associated with adaptive shifts in the ecology and morphology of species. The taxon cycle concept was explicitly formulated by biologist E. O. Wilson in 1961 after he surveyed the distributions, habitats, behavior and morphology of ant species in the Melanesian archipelago.
Priyanga Amarasekare is an American zoologist. She is a Professor of Ecology and Evolutionary Biology at the University of California, Los Angeles (UCLA), and distinguished fellow of the Ecological Society of America (ESA). Her research is in the fields of mathematical biology and trophic ecology, with a focus on understanding patterns of biodiversity, species dispersal and the impacts of climate change. She received a 2021 Guggenheim Fellowship and received ESA's Robert H. MacArthur Award in 2022.
References
- 1 2 Maino, James L.; Kearney, Michael R.; Nisbet, Roger M.; Kooijman, Sebastiaan A. L. M. (2014-01-01). "Reconciling theories for metabolic scaling". Journal of Animal Ecology. 83 (1): 20–29. doi: 10.1111/1365-2656.12085 . ISSN 1365-2656. PMID 23668377.
- 1 2 Kearney, Michael R.; White, Craig R. (2012-11-01). "Testing Metabolic Theories" (PDF). The American Naturalist. 180 (5): 546–565. doi:10.1086/667860. ISSN 0003-0147. PMID 23070317.
- ↑ van der Meer, Jaap (2006). "Metabolic theories in ecology". Trends in Ecology & Evolution. 21 (3): 136–140. doi:10.1016/j.tree.2005.11.004. ISSN 0169-5347. PMID 16701489.
- ↑ van der Meer, Jaap (2006). "An introduction to Dynamic Energy Budget (DEB) models have many more parameters and hence are more complicated - with special emphasis on multiple parameter estimations". Journal of Sea Research. 56 (2): 85–102. Bibcode:2006JSR....56...85V. doi:10.1016/j.seares.2006.03.001. ISSN 1385-1101.
- ↑ Kearney, Michael R.; White, Craig R. (2012-11-01). "Testing Metabolic Theories" (PDF). The American Naturalist. 180 (5): 546–565. doi:10.1086/667860. ISSN 0003-0147. PMID 23070317.
- ↑ White, Craig R.; Kearney, Michael R.; Matthews, Philip G. D.; Kooijman, Sebastiaan A. L. M.; Marshall, Dustin J. (2011-12-01). "A Manipulative Test of Competing Theories for Metabolic Scaling" (PDF). The American Naturalist. 178 (6): 746–754. doi:10.1086/662666. ISSN 0003-0147. PMID 22089869.
- ↑ Lika, Konstadia; Nisbet, Roger M. (2000-10-01). "A Dynamic Energy Budget model based on partitioning of net production". Journal of Mathematical Biology. 41 (4): 361–386. doi:10.1007/s002850000049. ISSN 0303-6812. PMID 11103872. S2CID 46678581.