Related Research Articles
An intron is any nucleotide sequence within a gene that is not expressed or operative in the final RNA product. The word intron is derived from the term intragenic region, i.e., a region inside a gene. The term intron refers to both the DNA sequence within a gene and the corresponding RNA sequence in RNA transcripts. The non-intron sequences that become joined by this RNA processing to form the mature RNA are called exons.

RNA splicing is a process in molecular biology where a newly-made precursor messenger RNA (pre-mRNA) transcript is transformed into a mature messenger RNA (mRNA). It works by removing all the introns and splicing back together exons. For nuclear-encoded genes, splicing occurs in the nucleus either during or immediately after transcription. For those eukaryotic genes that contain introns, splicing is usually needed to create an mRNA molecule that can be translated into protein. For many eukaryotic introns, splicing occurs in a series of reactions which are catalyzed by the spliceosome, a complex of small nuclear ribonucleoproteins (snRNPs). There exist self-splicing introns, that is, ribozymes that can catalyze their own excision from their parent RNA molecule. The process of transcription, splicing and translation is called gene expression, the central dogma of molecular biology.

MicroRNA (miRNA) are small, single-stranded, non-coding RNA molecules containing 21 to 23 nucleotides. Found in plants, animals and some viruses, miRNAs are involved in RNA silencing and post-transcriptional regulation of gene expression. miRNAs base-pair to complementary sequences in mRNA molecules, then silence said mRNA molecules by one or more of the following processes:
- Cleavage of the mRNA strand into two pieces,
- Destabilization of the mRNA by shortening its poly(A) tail, or
- Reducing translation of the mRNA into proteins.
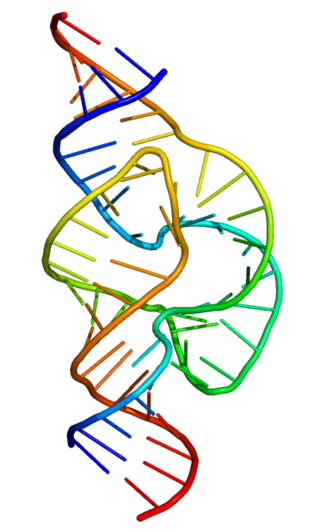
Ribozymes are RNA molecules that have the ability to catalyze specific biochemical reactions, including RNA splicing in gene expression, similar to the action of protein enzymes. The 1982 discovery of ribozymes demonstrated that RNA can be both genetic material and a biological catalyst, and contributed to the RNA world hypothesis, which suggests that RNA may have been important in the evolution of prebiotic self-replicating systems.
The RNA-induced silencing complex, or RISC, is a multiprotein complex, specifically a ribonucleoprotein, which functions in gene silencing via a variety of pathways at the transcriptional and translational levels. Using single-stranded RNA (ssRNA) fragments, such as microRNA (miRNA), or double-stranded small interfering RNA (siRNA), the complex functions as a key tool in gene regulation. The single strand of RNA acts as a template for RISC to recognize complementary messenger RNA (mRNA) transcript. Once found, one of the proteins in RISC, Argonaute, activates and cleaves the mRNA. This process is called RNA interference (RNAi) and it is found in many eukaryotes; it is a key process in defense against viral infections, as it is triggered by the presence of double-stranded RNA (dsRNA).

Drosha is a Class 2 ribonuclease III enzyme that in humans is encoded by the DROSHA gene. It is the primary nuclease that executes the initiation step of miRNA processing in the nucleus. It works closely with DGCR8 and in correlation with Dicer. It has been found significant in clinical knowledge for cancer prognosis and HIV-1 replication.
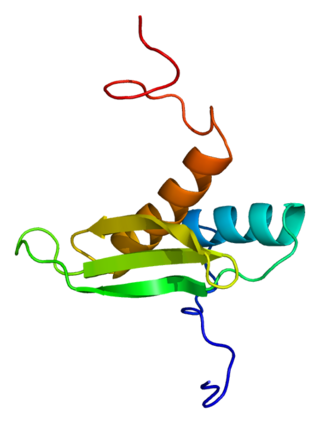
The microprocessor complex subunit DGCR8(DiGeorge syndrome critical region 8) is a protein that in humans is encoded by the DGCR8 gene. In other animals, particularly the common model organisms Drosophila melanogaster and Caenorhabditis elegans, the protein is known as Pasha. It is a required component of the RNA interference pathway.
RNA silencing or RNA interference refers to a family of gene silencing effects by which gene expression is negatively regulated by non-coding RNAs such as microRNAs. RNA silencing may also be defined as sequence-specific regulation of gene expression triggered by double-stranded RNA (dsRNA). RNA silencing mechanisms are conserved among most eukaryotes. The most common and well-studied example is RNA interference (RNAi), in which endogenously expressed microRNA (miRNA) or exogenously derived small interfering RNA (siRNA) induces the degradation of complementary messenger RNA. Other classes of small RNA have been identified, including piwi-interacting RNA (piRNA) and its subspecies repeat associated small interfering RNA (rasiRNA).
The Let-7 microRNA precursor was identified from a study of developmental timing in C. elegans, and was later shown to be part of a much larger class of non-coding RNAs termed microRNAs. miR-98 microRNA precursor from human is a let-7 family member. Let-7 miRNAs have now been predicted or experimentally confirmed in a wide range of species (MIPF0000002). miRNAs are initially transcribed in long transcripts called primary miRNAs (pri-miRNAs), which are processed in the nucleus by Drosha and Pasha to hairpin structures of about 70 nucleotide. These precursors (pre-miRNAs) are exported to the cytoplasm by exportin5, where they are subsequently processed by the enzyme Dicer to a ~22 nucleotide mature miRNA. The involvement of Dicer in miRNA processing demonstrates a relationship with the phenomenon of RNA interference.

The miR-124 microRNA precursor is a small non-coding RNA molecule that has been identified in flies, nematode worms, mouse and human. The mature ~21 nucleotide microRNAs are processed from hairpin precursor sequences by the Dicer enzyme, and in this case originates from the 3' arm. miR-124 has been found to be the most abundant microRNA expressed in neuronal cells. Experiments to alter expression of miR-124 in neural cells did not appear to affect differentiation. However these results are controversial since other reports have described a role for miR-124 during neuronal differentiation.

miR-196 is a non-coding RNA called a microRNA that has been shown to be expressed in humans and mice. miR-196 appears to be a vertebrate specific microRNA and has now been predicted or experimentally confirmed in a wide range of vertebrate species. In many species the miRNA appears to be expressed from intergenic regions in HOX gene clusters. The hairpin precursors are predicted based on base pairing and cross-species conservation—their extents are not known. In this case the mature sequence is excised from the 5' arm of the hairpin.

The mir-2 microRNA family includes the microRNA genes mir-2 and mir-13. Mir-2 is widespread in invertebrates, and it is the largest family of microRNAs in the model species Drosophila melanogaster. MicroRNAs from this family are produced from the 3' arm of the precursor hairpin. Leaman et al. showed that the miR-2 family regulates cell survival by translational repression of proapoptotic factors. Based on computational prediction of targets, a role in neural development and maintenance has been suggested.
MicroRNA sequencing (miRNA-seq), a type of RNA-Seq, is the use of next-generation sequencing or massively parallel high-throughput DNA sequencing to sequence microRNAs, also called miRNAs. miRNA-seq differs from other forms of RNA-seq in that input material is often enriched for small RNAs. miRNA-seq allows researchers to examine tissue-specific expression patterns, disease associations, and isoforms of miRNAs, and to discover previously uncharacterized miRNAs. Evidence that dysregulated miRNAs play a role in diseases such as cancer has positioned miRNA-seq to potentially become an important tool in the future for diagnostics and prognostics as costs continue to decrease. Like other miRNA profiling technologies, miRNA-Seq has both advantages and disadvantages.
In molecular biology mir-62 microRNA is a short RNA molecule. MicroRNAs function to regulate the expression levels of other genes by several mechanisms.
In molecular biology mir-241 microRNA is a short RNA molecule. MicroRNAs function to regulate the expression levels of other genes by several mechanisms.
David P. Bartel is an American molecular biologist best known for his work on microRNAs. Bartel is a Professor of Biology at the Massachusetts Institute of Technology, Member of the Whitehead Institute, and investigator of the Howard Hughes Medical Institute (HHMI).
In molecular biology, small nucleolar RNA derived microRNAs are microRNAs (miRNA) derived from small nucleolar RNA (snoRNA). MicroRNAs are usually derived from precursors known as pre-miRNAs, these pre-miRNAs are recognised and cleaved from a pri-miRNA precursor by the Pasha and Drosha proteins. However some microRNAs, mirtrons, are known to be derived from introns via a different pathway which bypasses Pasha and Drosha. Some microRNAs are also known to be derived from small nucleolar RNA.

Circular RNA is a type of single-stranded RNA which, unlike linear RNA, forms a covalently closed continuous loop. In circular RNA, the 3' and 5' ends normally present in an RNA molecule have been joined together. This feature confers numerous properties to circular RNA, many of which have only recently been identified.
In bioinformatics, TargetScan is a web server that predicts biological targets of microRNAs (miRNAs) by searching for the presence of sites that match the seed region of each miRNA. For many species, other types of sites, known as 3'-compensatory sites are also identified. These miRNA target predictions are regularly updated and improved by the laboratory of David Bartel in conjunction with the Whitehead Institute Bioinformatics and Research Computing Group.

The microprocessor complex is a protein complex involved in the early stages of processing microRNA (miRNA) and RNA interference (RNAi) in animal cells. The complex is minimally composed of the ribonuclease enzyme Drosha and the dimeric RNA-binding protein DGCR8, and cleaves primary miRNA substrates to pre-miRNA in the cell nucleus. Microprocessor is also the smaller of the two multi-protein complexes that contain human Drosha.
References
- ↑ Ruby, JG; Jan, CH; Bartel, DP (Jul 5, 2007). "Intronic microRNA precursors that bypass Drosha processing". Nature. 448 (7149): 83–6. Bibcode:2007Natur.448...83R. doi:10.1038/nature05983. PMC 2475599 . PMID 17589500.
- ↑ Wen, Jiayu; Ladewig, Erik; Shenker, Sol; Mohammed, Jaaved; Lai, Eric C. (2015). "Analysis of Nearly One Thousand Mammalian Mirtrons Reveals Novel Features of Dicer Substrates". PLOS Computational Biology. 11 (9): e1004441. Bibcode:2015PLSCB..11E4441W. doi: 10.1371/journal.pcbi.1004441 . PMC 4556696 . PMID 26325366.
- ↑ Ruby, JG; Jan, CH; Bartel, DP (Jul 5, 2007). "Intronic microRNA precursors that bypass Drosha processing". Nature. 448 (7149): 83–6. Bibcode:2007Natur.448...83R. doi:10.1038/nature05983. PMC 2475599 . PMID 17589500.
- ↑ Da Fonseca, Bruno Henrique Ribeiro; Domingues, Douglas Silva; Paschoal, Alexandre Rossi (2019-10-01). "mirtronDB: a mirtron knowledge base". Bioinformatics. 35 (19): 3873–3874. doi:10.1093/bioinformatics/btz153. ISSN 1367-4803. PMC 6761972 . PMID 30874795.
- ↑ Ruby, JG; Jan, CH; Bartel, DP (Jul 5, 2007). "Intronic microRNA precursors that bypass Drosha processing". Nature. 448 (7149): 83–6. Bibcode:2007Natur.448...83R. doi:10.1038/nature05983. PMC 2475599 . PMID 17589500.
- ↑ Jan, CH; Ruby, JG; Friedman, R; Bartel, DP (Jan 6, 2011). "Formation, regulation and evolution of Caenorhabditis elegans 3′UTRs" (PDF). Nature. 469 (7328): 97–101. Bibcode:2011Natur.469...97J. doi:10.1038/nature09616. PMC 3057491 . PMID 21085120.
- 1 2 Zhu, Q.-H.; Spriggs, A.; Matthew, L.; Fan, L.; Kennedy, G.; Gubler, F.; Helliwell, C. (30 July 2008). "A diverse set of microRNAs and microRNA-like small RNAs in developing rice grains". Genome Research. 18 (9): 1456–1465. doi:10.1101/gr.075572.107. PMC 2527712 . PMID 18687877.
- ↑ "mirtron". mirtron.gene-quantification.info. Retrieved 2022-12-10.
- ↑ Rorbach, Grzegorz; Unold, Olgierd; Konopka, Bogumil M. (2018). "Distinguishing mirtrons from canonical miRNAs with data exploration and machine learning methods". Scientific Reports. 8 (1): 7560. Bibcode:2018NatSR...8.7560R. doi:10.1038/s41598-018-25578-3. PMC 5953923 . PMID 29765080.
| | This molecular or cell biology article is a stub. You can help Wikipedia by expanding it. |