Ataxia is a neurological sign consisting of lack of voluntary coordination of muscle movements that can include gait abnormality, speech changes, and abnormalities in eye movements, that indicates dysfunction of parts of the nervous system that coordinate movement, such as the cerebellum.

The cerebellum is a major feature of the hindbrain of all vertebrates. Although usually smaller than the cerebrum, in some animals such as the mormyrid fishes it may be as large as it or even larger. In humans, the cerebellum plays an important role in motor control. It may also be involved in some cognitive functions such as attention and language as well as emotional control such as regulating fear and pleasure responses, but its movement-related functions are the most solidly established. The human cerebellum does not initiate movement, but contributes to coordination, precision, and accurate timing: it receives input from sensory systems of the spinal cord and from other parts of the brain, and integrates these inputs to fine-tune motor activity. Cerebellar damage produces disorders in fine movement, equilibrium, posture, and motor learning in humans.
Muscle memory is a form of procedural memory that involves consolidating a specific motor task into memory through repetition, which has been used synonymously with motor learning. When a movement is repeated over time, the brain creates a long-term muscle memory for that task, eventually allowing it to be performed with little to no conscious effort. This process decreases the need for attention and creates maximum efficiency within the motor and memory systems. Muscle memory is found in many everyday activities that become automatic and improve with practice, such as riding bikes, driving motor vehicles, playing ball sports, typing on keyboards, entering PINs, playing musical instruments, poker, martial arts, swimming, dancing, and drawing.

A gait is a manner of limb movements made during locomotion. Human gaits are the various ways in which humans can move, either naturally or as a result of specialized training. Human gait is defined as bipedal forward propulsion of the center of gravity of the human body, in which there are sinuous movements of different segments of the body with little energy spent. Varied gaits are characterized by differences such as limb movement patterns, overall velocity, forces, kinetic and potential energy cycles, and changes in contact with the ground.

The inferior olivary nucleus (ION), is a structure found in the medulla oblongata underneath the superior olivary nucleus. In vertebrates, the ION is known to coordinate signals from the spinal cord to the cerebellum to regulate motor coordination and learning. These connections have been shown to be tightly associated, as degeneration of either the cerebellum or the ION results in degeneration of the other.

Purkinje cells, or Purkinje neurons, are a class of GABAergic inhibitory neurons located in the cerebellum. They are named after their discoverer, Czech anatomist Jan Evangelista Purkyně, who characterized the cells in 1839.
The pedunculopontine nucleus (PPN) or pedunculopontine tegmental nucleus is a collection of neurons located in the upper pons in the brainstem. It lies caudal to the substantia nigra and adjacent to the superior cerebellar peduncle. It has two divisions of subnuclei; the pars compacta containing mainly cholinergic neurons, and the pars dissipata containing mainly glutamatergic neurons and some non-cholinergic neurons. The pedunculopontine nucleus is one of the main components of the reticular activating system. It was first described in 1909 by Louis Jacobsohn-Lask, a German neuroanatomist.
Dysmetria is a lack of coordination of movement typified by the undershoot or overshoot of intended position with the hand, arm, leg, or eye. It is a type of ataxia. It can also include an inability to judge distance or scale.
Central pattern generators (CPGs) are self-organizing biological neural circuits that produce rhythmic outputs in the absence of rhythmic input. They are the source of the tightly-coupled patterns of neural activity that drive rhythmic and stereotyped motor behaviors like walking, swimming, breathing, or chewing. The ability to function without input from higher brain areas still requires modulatory inputs, and their outputs are not fixed. Flexibility in response to sensory input is a fundamental quality of CPG-driven behavior. To be classified as a rhythmic generator, a CPG requires:
- "two or more processes that interact such that each process sequentially increases and decreases, and
- that, as a result of this interaction, the system repeatedly returns to its starting condition."
Motor control is the regulation of movements in organisms that possess a nervous system. Motor control includes conscious voluntary movements, subconscious muscle memory and involuntary reflexes, as well as instinctual taxis.
In physiology, motor coordination is the orchestrated movement of multiple body parts as required to accomplish intended actions, like walking. This coordination is achieved by adjusting kinematic and kinetic parameters associated with each body part involved in the intended movement. The modifications of these parameters typically relies on sensory feedback from one or more sensory modalities, such as proprioception and vision.

The posterior parietal cortex plays an important role in planned movements, spatial reasoning, and attention.
The neuroanatomy of memory encompasses a wide variety of anatomical structures in the brain.
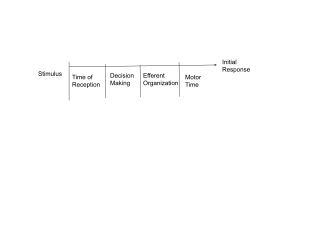
A motor program is an abstract metaphor of the central organization of movement and control of the many degrees of freedom involved in performing an action.p. 182 Signals transmitted through efferent and afferent pathways allow the central nervous system to anticipate, plan or guide movement. Evidence for the concept of motor programs include the following:p. 182

Central pattern generators are biological neural networks organized to produce any rhythmic output without requiring a rhythmic input. In mammals, locomotor CPGs are organized in the lumbar and cervical segments of the spinal cord, and are used to control rhythmic muscle output in the arms and legs. Certain areas of the brain initiate the descending neural pathways that ultimately control and modulate the CPG signals. In addition to this direct control, there exist different feedback loops that coordinate the limbs for efficient locomotion and allow for the switching of gaits under appropriate circumstances.
Gain field encoding is a hypothesis about the internal storage and processing of limb motion in the brain. In the motor areas of the brain, there are neurons which collectively have the ability to store information regarding both limb positioning and velocity in relation to both the body (intrinsic) and the individual's external environment (extrinsic). The input from these neurons is taken multiplicatively, forming what is referred to as a gain field. The gain field works as a collection of internal models off of which the body can base its movements. The process of encoding and recalling these models is the basis of muscle memory.

ATP/GTP binding protein 1 is gene that encodes the protein known as cytosolic carboxypeptidase 1 (CCP1), originally named NNA1. Mice with a naturally occurring mutation of the Agtpbp1 gene are known as pcd mice.

Megan Carey is a neuroscientist and Group Leader of the Neural Circuits and Behavior Laboratory at the Champalimaud Centre for the Unknown in Lisbon, Portugal. She is known for her work on how the cerebellum controls coordinated movement.
The study of animal locomotion is a branch of biology that investigates and quantifies how animals move.

Interlimb coordination is the coordination of the left and right limbs. It could be classified into two types of action: bimanual coordination and hands or feet coordination. Such coordination involves various parts of the nervous system and requires a sensory feedback mechanism for the neural control of the limbs. A model can be used to visualize the basic features, the control centre of locomotor movements, and the neural control of interlimb coordination. This coordination mechanism can be altered and adapted for better performance during locomotion in adults and for the development of motor skills in infants. The adaptive feature of interlimb coordination can also be applied to the treatment for CNS damage from stroke and the Parkinson's disease in the future.

