
A motor neuron is a neuron whose cell body is located in the motor cortex, brainstem or the spinal cord, and whose axon (fiber) projects to the spinal cord or outside of the spinal cord to directly or indirectly control effector organs, mainly muscles and glands. There are two types of motor neuron – upper motor neurons and lower motor neurons. Axons from upper motor neurons synapse onto interneurons in the spinal cord and occasionally directly onto lower motor neurons. The axons from the lower motor neurons are efferent nerve fibers that carry signals from the spinal cord to the effectors. Types of lower motor neurons are alpha motor neurons, beta motor neurons, and gamma motor neurons.

Skeletal muscle is one of the three types of vertebrate muscle tissue, the other being cardiac muscle and smooth muscle. They are part of the voluntary muscular system and typically are attached by tendons to bones of a skeleton. The skeletal muscle cells are much longer than in the other types of muscle tissue, and are also known as muscle fibers. The tissue of a skeletal muscle is striated – having a striped appearance due to the arrangement of the sarcomeres.
In biology, a motor unit is made up of a motor neuron and all of the skeletal muscle fibers innervated by the neuron's axon terminals, including the neuromuscular junctions between the neuron and the fibres. Groups of motor units often work together as a motor pool to coordinate the contractions of a single muscle. The concept was proposed by Charles Scott Sherrington.
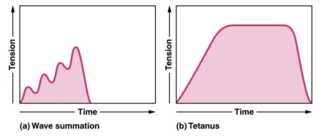
A tetanic contraction is a sustained muscle contraction evoked when the motor nerve that innervates a skeletal muscle emits action potentials at a very high rate. During this state, a motor unit has been maximally stimulated by its motor neuron and remains that way for some time. This occurs when a muscle's motor unit is stimulated by multiple impulses at a sufficiently high frequency. Each stimulus causes a twitch. If stimuli are delivered slowly enough, the tension in the muscle will relax between successive twitches. If stimuli are delivered at high frequency, the twitches will overlap, resulting in tetanic contraction. A tetanic contraction can be either unfused (incomplete) or fused (complete). An unfused tetanus is when the muscle fibers do not completely relax before the next stimulus because they are being stimulated at a fast rate; however there is a partial relaxation of the muscle fibers between the twitches. Fused tetanus is when there is no relaxation of the muscle fibers between stimuli and it occurs during a high rate of stimulation. A fused tetanic contraction is the strongest single-unit twitch in contraction. When tetanized, the contracting tension in the muscle remains constant in a steady state. This is the maximal possible contraction. During tetanic contractions, muscles can shorten, lengthen or remain constant length.

Muscle spindles are stretch receptors within the body of a skeletal muscle that primarily detect changes in the length of the muscle. They convey length information to the central nervous system via afferent nerve fibers. This information can be processed by the brain as proprioception. The responses of muscle spindles to changes in length also play an important role in regulating the contraction of muscles, for example, by activating motor neurons via the stretch reflex to resist muscle stretch.
Muscle fatigue is when muscles that were initially generating a normal amount of force, then experience a declining ability to generate force. It can be a result of vigorous exercise, but abnormal fatigue may be caused by barriers to or interference with the different stages of muscle contraction. There are two main causes of muscle fatigue: the limitations of a nerve’s ability to generate a sustained signal ; and the reduced ability of the muscle fiber to contract.

Muscle contraction is the activation of tension-generating sites within muscle cells. In physiology, muscle contraction does not necessarily mean muscle shortening because muscle tension can be produced without changes in muscle length, such as when holding something heavy in the same position. The termination of muscle contraction is followed by muscle relaxation, which is a return of the muscle fibers to their low tension-generating state.
Synaptogenesis is the formation of synapses between neurons in the nervous system. Although it occurs throughout a healthy person's lifespan, an explosion of synapse formation occurs during early brain development, known as exuberant synaptogenesis. Synaptogenesis is particularly important during an individual's critical period, during which there is a certain degree of synaptic pruning due to competition for neural growth factors by neurons and synapses. Processes that are not used, or inhibited during their critical period will fail to develop normally later on in life.
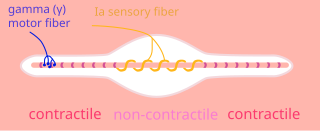
A gamma motor neuron, also called gamma motoneuron, or fusimotor neuron, is a type of lower motor neuron that takes part in the process of muscle contraction, and represents about 30% of (Aγ) fibers going to the muscle. Like alpha motor neurons, their cell bodies are located in the anterior grey column of the spinal cord. They receive input from the reticular formation of the pons in the brainstem. Their axons are smaller than those of the alpha motor neurons, with a diameter of only 5 μm. Unlike the alpha motor neurons, gamma motor neurons do not directly adjust the lengthening or shortening of muscles. However, their role is important in keeping muscle spindles taut, thereby allowing the continued firing of alpha neurons, leading to muscle contraction. These neurons also play a role in adjusting the sensitivity of muscle spindles.
Motor control is the regulation of movements in organisms that possess a nervous system. Motor control includes conscious voluntary movements, subconscious muscle memory and involuntary reflexes, as well as instinctual taxis.
Motor unit number estimation (MUNE) is a technique that uses electromyography to estimate the number of motor units in a muscle.

Muscle is a soft tissue, one of the four basic types of animal tissue. Muscle tissue gives skeletal muscles the ability to contract. Muscle is formed during embryonic development, in a process known as myogenesis. Muscle tissue contains special contractile proteins called actin and myosin which interact to cause movement. Among many other muscle proteins, present are two regulatory proteins, troponin and tropomyosin.
Myomeres are blocks of skeletal muscle tissue arranged in sequence, commonly found in aquatic chordates. Myomeres are separated from adjacent myomeres by connective fascia (myosepta) and most easily seen in larval fishes or in the olm. Myomere counts are sometimes used for identifying specimens, since their number corresponds to the number of vertebrae in the adults. Location varies, with some species containing these only near the tails, while some have them located near the scapular or pelvic girdles. Depending on the species, myomeres could be arranged in an epaxial or hypaxial manner. Hypaxial refers to ventral muscles and related structures while epaxial refers to more dorsal muscles. The horizontal septum divides these two regions in vertebrates from cyclostomes to gnathostomes. In terrestrial chordates, the myomeres become fused as well as indistinct, due to the disappearance of myosepta.
A motor pool consists of all individual motor neurons that innervate a single muscle. Each individual muscle fiber is innervated by only one motor neuron, but one motor neuron may innervate several muscle fibers. This distinction is physiologically significant because the size of a given motor pool determines the activity of the muscle it innervates: for example, muscles responsible for finer movements are innervated by motor pools consisting of higher numbers of individual motor neurons. Motor pools are also distinguished by the different classes of motor neurons that they contain. The size, composition, and anatomical location of each motor pool is tightly controlled by complex developmental pathways.
Beta motor neurons, also called beta motoneurons, are a few kind of lower motor neuron, along with alpha motor neurons and gamma motor neurons. Beta motor neurons innervate intrafusal fibers of muscle spindles with collaterals to extrafusal fibers - a type of slow twitch fiber. Also, axons of alpha, beta, and gamma motor neurons become myelinated. Moreover, these efferent neurons originate from the anterior grey column of the spinal cord and travel to skeletal muscles. However, the larger diameter alpha motor fibers require higher conduction velocity than beta and gamma.
The motor unit consists of a voluntary alpha motoneuron and all of the collective muscle fibers that it controls, known as the effector muscle. The alpha motoneuron communicates with acetylcholine receptors on the motor end plate of the effector muscle. Reception of acetylcholine neurotransmitters on the motor end plate causes contraction of that effector muscle.
Normal aging movement control in humans is about the changes in the muscles, motor neurons, nerves, sensory functions, gait, fatigue, visual and manual responses, in men and women as they get older but who do not have neurological, muscular or neuromuscular disorder. With aging, neuromuscular movements are impaired, though with training or practice, some aspects may be prevented.
Henneman’s size principle describes relationships between properties of motor neurons and the muscle fibers they innervate and thus control, which together are called motor units. Motor neurons with large cell bodies tend to innervate fast-twitch, high-force, less fatigue-resistant muscle fibers, whereas motor neurons with small cell bodies tend to innervate slow-twitch, low-force, fatigue-resistant muscle fibers. In order to contract a particular muscle, motor neurons with small cell bodies are recruited before motor neurons with large cell bodies. It was proposed by Elwood Henneman.

James B. Preston was an American neurophysiologist at State University of New York Upstate Medical University whose research was fundamental to discovering how our brains control movement. Over the course of his career, he published over forty research based articles in his field. Preston was the chairman of numerous national committees and former President of the Association of Chairs of Departments of Physiology.
Elwood Henneman was an American neurophysiologist who studied the properties of vertebrate motor neurons.

