
The visual cortex of the brain is the area of the cerebral cortex that processes visual information. It is located in the occipital lobe. Sensory input originating from the eyes travels through the lateral geniculate nucleus in the thalamus and then reaches the visual cortex. The area of the visual cortex that receives the sensory input from the lateral geniculate nucleus is the primary visual cortex, also known as visual area 1 (V1), Brodmann area 17, or the striate cortex. The extrastriate areas consist of visual areas 2, 3, 4, and 5.

The visual system comprises the sensory organ and parts of the central nervous system which gives organisms the sense of sight as well as enabling the formation of several non-image photo response functions. It detects and interprets information from the optical spectrum perceptible to that species to "build a representation" of the surrounding environment. The visual system carries out a number of complex tasks, including the reception of light and the formation of monocular neural representations, colour vision, the neural mechanisms underlying stereopsis and assessment of distances to and between objects, the identification of particular object of interest, motion perception, the analysis and integration of visual information, pattern recognition, accurate motor coordination under visual guidance, and more. The neuropsychological side of visual information processing is known as visual perception, an abnormality of which is called visual impairment, and a complete absence of which is called blindness. Non-image forming visual functions, independent of visual perception, include the pupillary light reflex (PLR) and circadian photoentrainment.

Torsten Nils Wiesel is a Swedish neurophysiologist. Together with David H. Hubel, he received the 1981 Nobel Prize in Physiology or Medicine, for their discoveries concerning information processing in the visual system; the prize was shared with Roger W. Sperry for his independent research on the cerebral hemispheres.

David Hunter Hubel was a Canadian American neurophysiologist noted for his studies of the structure and function of the visual cortex. He was co-recipient with Torsten Wiesel of the 1981 Nobel Prize in Physiology or Medicine, for their discoveries concerning information processing in the visual system. For much of his career, Hubel was the John Franklin Enders University Professor of Neurobiology at Harvard Medical School. In 1978, Hubel and Wiesel were awarded the Louisa Gross Horwitz Prize from Columbia University.In 1983, Hubel received the Golden Plate Award of the American Academy of Achievement.
A cortical column, also called hypercolumn, macrocolumn, functional column or sometimes cortical module, is a group of neurons in the cortex of the brain that can be successively penetrated by a probe inserted perpendicularly to the cortical surface, and which have nearly identical receptive fields. Neurons within a minicolumn (microcolumn) encode similar features, whereas a hypercolumn "denotes a unit containing a full set of values for any given set of receptive field parameters". A cortical module is defined as either synonymous with a hypercolumn (Mountcastle) or as a tissue block of multiple overlapping hypercolumns.
According to Alonso and Chen (2008),
The receptive field is a portion of sensory space that can elicit neuronal responses when stimulated. The sensory space can be defined in a single dimension, two dimensions or multiple dimensions. The neuronal response can be defined as firing rate or include also subthreshold activity.
Neuronal tuning refers to the hypothesized property of brain cells by which they selectively represent a particular type of sensory, association, motor, or cognitive information. Some neuronal responses have been hypothesized to be optimally tuned to specific patterns through experience. Neuronal tuning can be strong and sharp, as observed in primary visual cortex, or weak and broad, as observed in neural ensembles. Single neurons are hypothesized to be simultaneously tuned to several modalities, such as visual, auditory, and olfactory. Neurons hypothesized to be tuned to different signals are often hypothesized to integrate information from the different sources. In computational models called neural networks, such integration is the major principle of operation. The best examples of neuronal tuning can be seen in the visual, auditory, olfactory, somatosensory, and memory systems, although due to the small number of stimuli tested the generality of neuronal tuning claims is still an open question.
Ocular dominance columns are stripes of neurons in the visual cortex of certain mammals that respond preferentially to input from one eye or the other. The columns span multiple cortical layers, and are laid out in a striped pattern across the surface of the striate cortex (V1). The stripes lie perpendicular to the orientation columns.
The neocognitron is a hierarchical, multilayered artificial neural network proposed by Kunihiko Fukushima in 1979. It has been used for handwritten character recognition and other pattern recognition tasks, and served as the inspiration for convolutional neural networks.
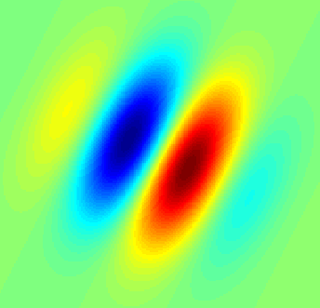
A simple cell in the primary visual cortex is a cell that responds primarily to oriented edges and gratings. These cells were discovered by Torsten Wiesel and David Hubel in the late 1950s.
Complex cells can be found in the primary visual cortex (V1), the secondary visual cortex (V2), and Brodmann area 19 (V3).

The colour centre is a region in the brain primarily responsible for visual perception and cortical processing of colour signals received by the eye, which ultimately results in colour vision. The colour centre in humans is thought to be located in the ventral occipital lobe as part of the visual system, in addition to other areas responsible for recognizing and processing specific visual stimuli, such as faces, words, and objects. Many functional magnetic resonance imaging (fMRI) studies in both humans and macaque monkeys have shown colour stimuli to activate multiple areas in the brain, including the fusiform gyrus and the lingual gyrus. These areas, as well as others identified as having a role in colour vision processing, are collectively labelled visual area 4 (V4). The exact mechanisms, location, and function of V4 are still being investigated.
A hypercomplex cell is a type of visual processing neuron in the mammalian cerebral cortex. Initially discovered by David Hubel and Torsten Wiesel in 1965, hypercomplex cells are defined by the property of end-stopping, which is a decrease in firing strength with increasingly larger stimuli. The sensitivity to stimulus length is accompanied by selectivity for the specific orientation, motion, and direction of stimuli. For example, a hypercomplex cell may only respond to a line at 45˚ that travels upward. Elongating the line would result in a proportionately weaker response. Ultimately, hypercomplex cells can provide a means for the brain to visually perceive corners and curves in the environment by identifying the ends of a given stimulus.
Feature detection is a process by which the nervous system sorts or filters complex natural stimuli in order to extract behaviorally relevant cues that have a high probability of being associated with important objects or organisms in their environment, as opposed to irrelevant background or noise.
Monocular deprivation is an experimental technique used by neuroscientists to study central nervous system plasticity. Generally, one of an animal's eyes is sutured shut during a period of high cortical plasticity. This manipulation serves as an animal model for amblyopia, a permanent deficit in visual sensation not due to abnormalities in the eye.
Oblique effect is the name given to the relative deficiency in perceptual performance for oblique contours as compared to the performance for horizontal or vertical contours.

Due to the effect of a spatial context or temporal context, the perceived orientation of a test line or grating pattern can appear tilted away from its physical orientation. The tilt illusion (TI) is the phenomenon that the perceived orientation of a test line or grating is altered by the presence of surrounding lines or grating with a different orientation. And the tilt aftereffect (TAE) is the phenomenon that the perceived orientation is changed after prolonged inspection of another oriented line or grating.
Binocular neurons are neurons in the visual system that assist in the creation of stereopsis from binocular disparity. They have been found in the primary visual cortex where the initial stage of binocular convergence begins. Binocular neurons receive inputs from both the right and left eyes and integrate the signals together to create a perception of depth.

Russell L. De Valois was an American scientist recognized for his pioneering research on spatial and color vision.
Orientation selectivity is expressed by cells within the visual cortex, when such cells increase impulse or signal activity for specific oriented degree of shape presented within the visual field. Orientation selectivity can also be expressed by simple cells if the orientation of a stimulus is orthogonal to the preferred degree of orientation, which results in the inhibition of impulse activity.


