
Amniotes are tetrapod vertebrate animals belonging to the clade Amniota, a large group that comprises the vast majority of living terrestrial and semiaquatic vertebrates. Amniotes evolved from amphibian ancestors during the Carboniferous period and further diverged into two groups, namely the sauropsids and synapsids. They are distinguished from the other living tetrapod clade — the non-amniote lissamphibians — by the development of three extraembryonic membranes, thicker and keratinized skin, and costal respiration. Additional unique features are the presence of adrenocortical and chromaffin tissues as a discrete pair of glands near their kidneys, which are more complex, the presence of an astragalus for better extremity range of motion, and the complete loss of metamorphosis, gill and skin breathing, and any lateral line system.

Ichthyostega is an extinct genus of limbed tetrapodomorphs from the Late Devonian of what is now Greenland. It was among the earliest four-limbed vertebrates ever in the fossil record and was one of the first with weight-bearing adaptations for terrestrial locomotion. Ichthyostega possessed lungs and limbs that helped it navigate through shallow water in swamps. Although Ichthyostega is often labelled a 'tetrapod' because of its limbs and fingers, it evolved long before true crown group tetrapods and could more accurately be referred to as a stegocephalian or stem tetrapod. Likewise, while undoubtedly of amphibian build and habit, it is not a true member of the group in the narrow sense, as the first modern amphibians appeared in the Triassic Period. Until finds of other early stegocephalians and closely related fishes in the late 20th century, Ichthyostega stood alone as a transitional fossil between fish and tetrapods, combining fish and tetrapod features. Newer research has shown that it had an unusual anatomy, functioning more akin to a seal than a salamander, as previously assumed.

"Labyrinthodontia" is an informal grouping of extinct predatory amphibians which were major components of ecosystems in the late Paleozoic and early Mesozoic eras. Traditionally considered a subclass of the class Amphibia, modern classification systems recognize that labyrinthodonts are not a formal natural group (clade) exclusive of other tetrapods. Instead, they consistute an evolutionary grade, ancestral to living tetrapods such as lissamphibians and amniotes. "Labyrinthodont"-grade vertebrates evolved from lobe-finned fishes in the Devonian, though a formal boundary between fish and amphibian is difficult to define at this point in time.

Reptiliomorpha is a clade containing the amniotes and those tetrapods that share a more recent common ancestor with amniotes than with living amphibians (lissamphibians). It was defined by Michel Laurin (2001) and Vallin and Laurin (2004) as the largest clade that includes Homo sapiens, but not Ascaphus truei. Laurin and Reisz (2020) defined Pan-Amniota as the largest total clade containing Homo sapiens, but not Pipa pipa, Caecilia tentaculata, and Siren lacertina.

Diadectes is an extinct genus of large reptiliomorphs or synapsids that lived during the early Permian period. Diadectes was one of the first herbivorous tetrapods, and also one of the first fully terrestrial vertebrates to attain large size.

Temnospondyli or temnospondyls is a diverse ancient order of small to giant tetrapods—often considered primitive amphibians—that flourished worldwide during the Carboniferous, Permian and Triassic periods, with fossils being found on every continent. A few species continued into the Jurassic and Early Cretaceous periods, but all had gone extinct by the Late Cretaceous. During about 210 million years of evolutionary history, they adapted to a wide range of habitats, including freshwater, terrestrial, and even coastal marine environments. Their life history is well understood, with fossils known from the larval stage, metamorphosis and maturity. Most temnospondyls were semiaquatic, although some were almost fully terrestrial, returning to the water only to breed. These temnospondyls were some of the first vertebrates fully adapted to life on land. Although temnospondyls are amphibians, many had characteristics such as scales and large armour-like bony plates (osteoderms) that generally distinguish them from the modern soft-bodied lissamphibians.

Seymouria is an extinct genus of seymouriamorph from the Early Permian of North America and Europe. Although they were amphibians, Seymouria were well-adapted to life on land, with many reptilian features—so many, in fact, that Seymouria was first thought to be a primitive reptile. It is primarily known from two species, Seymouria baylorensis and Seymouria sanjuanensis. The type species, S. baylorensis, is more robust and specialized, though its fossils have only been found in Texas. On the other hand, Seymouria sanjuanensis is more abundant and widespread. This smaller species is known from multiple well-preserved fossils, including a block of six skeletons found in the Cutler Formation of New Mexico, and a pair of fully grown skeletons from the Tambach Formation of Germany, which were fossilized lying next to each other.
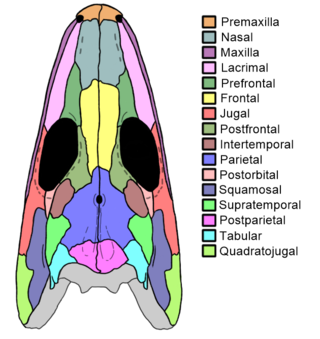
Proterogyrinus is an extinct genus of early tetrapods from the order Embolomeri. Fossil remains of Proterogyrinus have been found in Scotland, UK, and West Virginia, United States, and date back to the Serpukhovian, which is from about 331 to 323 million years ago. The genus was originally named by renowned vertebrate paleontologist Alfred Sherwood Romer in 1970. A comprehensive redescription was later published by Canadian paleontologist Robert Holmes in 1984. The generic name "Proterogyrinus" is Greek for "earlier wanderer" or "earlier tadpole". This name was chosen by Romer in keeping with a trend of naming long-bodied early tetrapods with the suffix "-gyrinus".

Crassigyrinus is an extinct genus of carnivorous stem tetrapod from the Early Carboniferous Limestone Coal Group of Scotland and possibly Greer, West Virginia.

Solenodonsaurus is an extinct genus of reptiliomorphs that lived in what is now Czech Republic, during the Westphalian stage.

Colosteidae is a family of stegocephalians that lived in the Carboniferous period. They possessed a variety of characteristics from different tetrapod or stem-tetrapod groups, which made them historically difficult to classify. They are now considered to be part of a lineage intermediate between the earliest Devonian terrestrial vertebrates, and the different groups ancestral to all modern tetrapods, such as temnospondyls and reptiliomorphs.

The hyomandibula, commonly referred to as hyomandibular [bone], is a set of bones that is found in the hyoid region in most fishes. It usually plays a role in suspending the jaws and/or operculum. It is commonly suggested that in tetrapods, the hyomandibula evolved into the columella (stapes).

Rhinesuchidae is a family of tetrapods that lived primarily in the Permian period. They belonged to the broad group Temnospondyli, a successful and diverse collection of semiaquatic tetrapods which modern amphibians are probably descended from. Rhinesuchids can be differentiated from other temnospondyls by details of their skulls, most notably the interior structure of their otic notches at the back of the skull. They were among the earliest-diverging members of the Stereospondyli, a subgroup of temnospondyls with flat heads and aquatic habits. Although more advanced stereospondyls evolved to reach worldwide distribution in the Triassic period, rhinesuchids primarily lived in the high-latitude environments of Gondwana during the Guadalupian and Lopingian epochs of the Permian. The taxonomy of this family has been convoluted, with more than twenty species having been named in the past; a 2017 review recognized only eight of them to be valid. While several purported members of this group have been reported to have lived in the Triassic period, most are either dubious or do not belong to the group. However, at least one valid genus of rhinesuchid is known from the early Triassic, a small member known as Broomistega. The most recent formal definition of Rhinesuchidae, advocated by Mariscano et al. (2017) is that of a stem-based clade containing all taxa more closely related to Rhinesuchus whaitsi than to Lydekkerina huxleyi or Peltobatrachus pustulatus. A similar alternate definition is that Rhinesuchidae is a stem-based clade containing all taxa more closely related to Uranocentrodon senekalensis than to Lydekkerina huxleyi, Trematosaurus brauni, or Mastodonsaurus giganteus.
Doleserpeton is an extinct, monospecific genus of dissorophoidean temnospondyl within the family Amphibamidae that lived during the Upper Permian, 285 million years ago. Doleserpeton is represented by a single species, Doleserpeton annectens, which was first described by John R. Bolt in 1969. Fossil evidence of Doleserpeton was recovered from the Dolese Brothers Limestone Quarry in Fort Sill, Oklahoma. The genus name Doleserpeton is derived from the initial discovery site in Dolese quarry of Oklahoma and the Greek root "herp-", meaning "low or close to the ground". This transitional fossil displays primitive traits of amphibians that allowed for successful adaptation from aquatic to terrestrial environments. In many phylogenies, lissamphibians appear as the sister group of Doleserpeton.

Dissorophus (DI-soh-ROH-fus) is an extinct genus of temnospondyl amphibian that lived during the Early Permian Period about 273 million years ago. Its fossils have been found in Texas and in Oklahoma in North America. Its heavy armor and robust build indicate Dissorophus was active on land, similar to other members of the clade Dissorophidae that are known from the Late Carboniferous to the Early Permian periods. Dissorphus is distinguished by its small body size, disproportionately large head and short trunk.

Lapillopsis is an extinct genus of stereospondyl temnospondyl within the family Lapillopsidae. Fossils belonging to the genus have been found in the Arcadia Formation of Queensland, Australia.
Perryella is an extinct genus of dvinosaurian(?) temnospondyl from the Permian of Oklahoma.
Reiszerpeton is an extinct genus of dissorophid temnospondyl known from the Early Permian Archer City Formation of Texas. It is known solely from the holotype, MCZ 1911, a complete skull. This specimen was originally referred to the amphibamiform Tersomius texensis. A reappraisal of the holotype of T. texensis and a number of other referred specimens by Maddin et al. (2013) noted a number of differences from both T. texensis and amphibamiforms more broadly that suggested affinities with the Dissorophidae. This was confirmed by a phylogenetic analysis, which placed it as the sister taxon to the Eucacopinae. Reiszerpeton is known only from the type species, R. renascentis, which was named for Canadian paleontologist Robert Reisz. The species name refers to the recognition of Reisz as a "renaissance paleontologist." It is differentiated from other dissorophids by its small size, small and more numerous maxillary teeth, smooth cranial ornamentation, and greater distance between the orbit and the otic notch.
Innovations conventionally associated with terrestrially first appeared in aquatic elpistostegalians such as Panderichthys rhombolepis, Elpistostege watsoni, and Tiktaalik roseae. Phylogenetic analyses distribute the features that developed along the tetrapod stem and display a stepwise process of character acquisition, rather than abrupt. The complete transition occurred over a period of 30 million years beginning with the tetrapodomorph diversification in the Middle Devonian.

Spiracles are openings on the surface of some animals, which usually lead to respiratory systems.

