Related Research Articles

In cell biology a centriole is a cylindrical organelle composed mainly of a protein called tubulin. Centrioles are found in most eukaryotic cells, but are not present in conifers (Pinophyta), flowering plants (angiosperms) and most fungi, and are only present in the male gametes of charophytes, bryophytes, seedless vascular plants, cycads, and Ginkgo. A bound pair of centrioles, surrounded by a highly ordered mass of dense material, called the pericentriolar material (PCM), makes up a structure called a centrosome.

A spermatozoon is a motile sperm cell produced by male animals relying on internal fertilization. A spermatozoon is a moving form of the haploid cell that is the male gamete that joins with an ovum to form a zygote.

The cilium is a short hair-like membrane protrusion from many types of eukaryotic cell. The cilium has the shape of a slender threadlike projection that extends from the surface of the much larger cell body. Eukaryotic flagella found on sperm cells and many protozoans have a similar structure to motile cilia that enables swimming through liquids; they are longer than cilia and have a different undulating motion.
Fertilisation or fertilization, also known as generative fertilisation, syngamy and impregnation, is the fusion of gametes to give rise to a zygote and initiate its development into a new individual organism or offspring. While processes such as insemination or pollination, which happen before the fusion of gametes, are also sometimes informally referred to as fertilisation, these are technically separate processes. The cycle of fertilisation and development of new individuals is called sexual reproduction. During double fertilisation in angiosperms, the haploid male gamete combines with two haploid polar nuclei to form a triploid primary endosperm nucleus by the process of vegetative fertilisation.
In cell biology, the centrosome is an organelle that serves as the main microtubule organizing center (MTOC) of the animal cell, as well as a regulator of cell-cycle progression. The centrosome provides structure for the cell. The centrosome is thought to have evolved only in the metazoan lineage of eukaryotic cells. Fungi and plants lack centrosomes and therefore use other structures to organize their microtubules. Although the centrosome has a key role in efficient mitosis in animal cells, it is not essential in certain fly and flatworm species.

Spermatogenesis is the process by which haploid spermatozoa develop from germ cells in the seminiferous tubules of the testicle. This process starts with the mitotic division of the stem cells located close to the basement membrane of the tubules. These cells are called spermatogonial stem cells. The mitotic division of these produces two types of cells. Type A cells replenish the stem cells, and type B cells differentiate into primary spermatocytes. The primary spermatocyte divides meiotically into two secondary spermatocytes; each secondary spermatocyte divides into two equal haploid spermatids by Meiosis II. The spermatids are transformed into spermatozoa (sperm) by the process of spermiogenesis. These develop into mature spermatozoa, also known as sperm cells. Thus, the primary spermatocyte gives rise to two cells, the secondary spermatocytes, and the two secondary spermatocytes by their subdivision produce four spermatozoa and four haploid cells.
An oocyte, oöcyte, or ovocyte is a female gametocyte or germ cell involved in reproduction. In other words, it is an immature ovum, or egg cell. An oocyte is produced in a female fetus in the ovary during female gametogenesis. The female germ cells produce a primordial germ cell (PGC), which then undergoes mitosis, forming oogonia. During oogenesis, the oogonia become primary oocytes. An oocyte is a form of genetic material that can be collected for cryoconservation.

Karyogamy is the final step in the process of fusing together two haploid eukaryotic cells, and refers specifically to the fusion of the two nuclei. Before karyogamy, each haploid cell has one complete copy of the organism's genome. In order for karyogamy to occur, the cell membrane and cytoplasm of each cell must fuse with the other in a process known as plasmogamy. Once within the joined cell membrane, the nuclei are referred to as pronuclei. Once the cell membranes, cytoplasm, and pronuclei fuse, the resulting single cell is diploid, containing two copies of the genome. This diploid cell, called a zygote or zygospore can then enter meiosis, or continue to divide by mitosis. Mammalian fertilization uses a comparable process to combine haploid sperm and egg cells (gametes) to create a diploid fertilized egg.

A pronucleus denotes the nucleus found in either a sperm or egg cell during the process of fertilization. The sperm cell undergoes a transformation into a pronucleus after entering the egg cell but prior to the fusion of the genetic material of both the sperm and egg. In contrast, the egg cell possesses a pronucleus once it becomes haploid, not upon the arrival of the sperm cell. Haploid cells, such as sperm and egg cells in humans, carry half the number of chromosomes present in somatic cells, with 23 chromosomes compared to the 46 found in somatic cells. It is noteworthy that the male and female pronuclei do not physically merge, although their genetic material does. Instead, their membranes dissolve, eliminating any barriers between the male and female chromosomes, facilitating the combination of their chromosomes into a single diploid nucleus in the resulting embryo, which contains a complete set of 46 chromosomes.
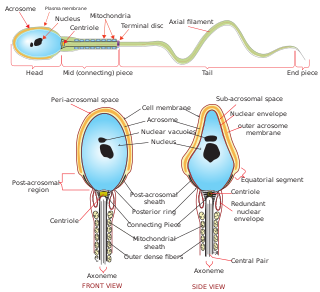
Sperm is the male reproductive cell, or gamete, in anisogamous forms of sexual reproduction. Animals produce motile sperm with a tail known as a flagellum, which are known as spermatozoa, while some red algae and fungi produce non-motile sperm cells, known as spermatia. Flowering plants contain non-motile sperm inside pollen, while some more basal plants like ferns and some gymnosperms have motile sperm.
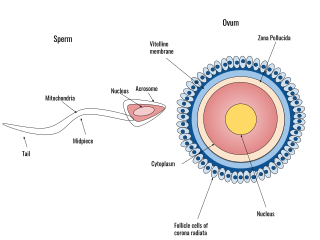
Human fertilization is the union of an egg and sperm, occurring primarily in the ampulla of the fallopian tube. The result of this union leads to the production of a fertilized egg called a zygote, initiating embryonic development. Scientists discovered the dynamics of human fertilization in the 19th century.

A polar body is a small haploid cell that is formed at the same time as an egg cell during oogenesis, but generally does not have the ability to be fertilized. It is named from its polar position in the egg.
An asymmetric cell division produces two daughter cells with different cellular fates. This is in contrast to symmetric cell divisions which give rise to daughter cells of equivalent fates. Notably, stem cells divide asymmetrically to give rise to two distinct daughter cells: one copy of the original stem cell as well as a second daughter programmed to differentiate into a non-stem cell fate.

Sperm motility describes the ability of sperm to move properly through the female reproductive tract or through water to reach the egg. Sperm motility can also be thought of as the quality, which is a factor in successful conception; sperm that do not "swim" properly will not reach the egg in order to fertilize it. Sperm motility in mammals also facilitates the passage of the sperm through the cumulus oophorus and the zona pellucida, which surround the mammalian oocyte.

Centrins, also known as caltractins, are a family of calcium-binding phosphoproteins found in the centrosome of eukaryotes. Centrins are small calcium binding proteins that are ubiquitous centrosome components. There are about 350 “signature” proteins that are unique to eukaryotic cells but have no significant homology to proteins in archaea and bacteria. They are a type of protein that is essential and present in almost all eukaryotic cells and are found in the centrioles and pericentriolar lattice. Human centrin genes are CETN1, CETN2 and CETN3.

Centromere protein J is a protein that in humans is encoded by the CENPJ gene. It is also known as centrosomal P4.1-associated protein (CPAP). During cell division, this protein plays a structural role in the maintenance of centrosome integrity and normal spindle morphology, and it is involved in microtubule disassembly at the centrosome. This protein can function as a transcriptional coactivator in the Stat5 signaling pathway and also as a coactivator of NF-kappaB-mediated transcription, likely via its interaction with the coactivator p300/CREB-binding protein.

The fallopian tubes, also known as uterine tubes, oviducts or salpinges, are paired tubular sex organs in the human female body that stretch from the ovaries to the uterus. The fallopian tubes are part of the female reproductive system. In other vertebrates, they are only called oviducts.

Pericentriolar material is a highly structured, dense mass of protein which makes up the part of the animal centrosome that surrounds the two centrioles. The PCM contains proteins responsible for microtubule nucleation and anchoring including γ-tubulin, pericentrin and ninein.

Centrosomal protein of 152 kDa, also known as Cep152, is a protein that in humans is encoded by the CEP152 gene. It is the ortholog of the Drosophila melanogaster gene asterless (asl) and both are required for centriole duplication.

Centrosomes are the major microtubule organizing centers (MTOC) in mammalian cells. Failure of centrosome regulation can cause mistakes in chromosome segregation and is associated with aneuploidy. A centrosome is composed of two orthogonal cylindrical protein assemblies, called centrioles, which are surrounded by a protein dense amorphous cloud of pericentriolar material (PCM). The PCM is essential for nucleation and organization of microtubules. The centrosome cycle is important to ensure that daughter cells receive a centrosome after cell division. As the cell cycle progresses, the centrosome undergoes a series of morphological and functional changes. Initiation of the centrosome cycle occurs early in the cell cycle in order to have two centrosomes by the time mitosis occurs.
References
- 1 2 3 Blachon, S; Cai, X; Roberts, K. A; Yang, K; Polyanovsky, A; Church, A; Avidor-Reiss, T (2009). "A Proximal Centriole-Like Structure is Present in Drosophila Spermatids and Can Serve as a Model to Study Centriole Duplication". Genetics. 182 (1): 133–44. doi:10.1534/genetics.109.101709. PMC 2674812 . PMID 19293139.
- 1 2 Khire, Atul; Jo, Kyoung H; Kong, Dong; Akhshi, Tara; Blachon, Stephanie; Cekic, Anthony R; Hynek, Sarah; Ha, Andrew; Loncarek, Jadranka; Mennella, Vito; Avidor-Reiss, Tomer (2016). "Centriole Remodeling during Spermiogenesis in Drosophila". Current Biology. 26 (23): 3183–3189. doi:10.1016/j.cub.2016.07.006. PMC 5245371 . PMID 28094036.
- ↑ Blachon, Stephanie; Khire, Atul; Avidor-Reiss, Tomer (2014). "The Origin of the Second Centriole in the Zygote of Drosophila melanogaster". Genetics. 197 (1): 199–205. doi:10.1534/genetics.113.160523. PMC 4012480 . PMID 24532732.
- ↑ Fishman, E. L; Jo, Kyoung; Ha, Andrew; Royfman, Rachel; Zinn, Ashtyn; Krishnamurthy, Malathi; Avidor-Reiss, Tomer (2017). "Atypical centrioles are present in Tribolium sperm". Open Biology. 7 (3): 160334. doi:10.1098/rsob.160334. PMC 5376708 . PMID 28298310.
- 1 2 Fishman, Emily L; Jo, Kyoung; Nguyen, Quynh P. H; Kong, Dong; Royfman, Rachel; Cekic, Anthony R; Khanal, Sushil; Miller, Ann L; Simerly, Calvin; Schatten, Gerald; Loncarek, Jadranka; Mennella, Vito; Avidor-Reiss, Tomer (2018). "A novel atypical sperm centriole is functional during human fertilization". Nature Communications. 9 (1): 2210. doi:10.1038/s41467-018-04678-8. PMC 5992222 . PMID 29880810.
- ↑ Khanal, S., M.R. Leung, A. Royfman, E.L. Fishman, B. Saltzman, H. Bloomfield-Gadelha, T. Zeev-Ben-Mordehai, and T. Avidor-Reiss. 2021. A dynamic basal complex modulates mammalian sperm movement. Nat Commun. 12:3808.. https://doi.org/10.1038/s41467-021-24011-0