
A cell wall is a structural layer surrounding some types of cells, just outside the cell membrane. It can be tough, flexible, and sometimes rigid. It provides the cell with both structural support and protection, and also acts as a filtering mechanism. Cell walls are absent in many eukaryotes, including animals, but are present in some other ones like fungi, algae and plants, and in most prokaryotes. A major function is to act as pressure vessels, preventing over-expansion of the cell when water enters.

Plant cells are the cells present in green plants, photosynthetic eukaryotes of the kingdom Plantae. Their distinctive features include primary cell walls containing cellulose, hemicelluloses and pectin, the presence of plastids with the capability to perform photosynthesis and store starch, a large vacuole that regulates turgor pressure, the absence of flagella or centrioles, except in the gametes, and a unique method of cell division involving the formation of a cell plate or phragmoplast that separates the new daughter cells.

Phloem is the living tissue in vascular plants that transports the soluble organic compounds made during photosynthesis and known as photosynthates, in particular the sugar sucrose, to the rest of the plant. This transport process is called translocation. In trees, the phloem is the innermost layer of the bark, hence the name, derived from the Ancient Greek word φλοιός (phloiós), meaning "bark". The term was introduced by Carl Nägeli in 1858.

Root pressure is the transverse osmotic pressure within the cells of a root system that causes sap to rise through a plant stem to the leaves.
Protoplasm is the living part of a cell that is surrounded by a plasma membrane. It is a mixture of small molecules such as ions, monosaccharides, amino acids, and macromolecules such as proteins, polysaccharides, lipids, etc.

The endodermis is the innermost layer of cortex in land plants. It is a cylinder of compact living cells, the radial walls of which are impregnated with hydrophobic substances to restrict apoplastic flow of water to the inside. The endodermis is the boundary between the cortex and the stele.

Suberin, cutin and lignins are complex, higher plant epidermis and periderm cell-wall macromolecules, forming a protective barrier. Suberin, a complex polyester biopolymer, is lipophilic, and composed of long chain fatty acids called suberin acids, and glycerol. Suberins and lignins are considered covalently linked to lipids and carbohydrates, respectively, and lignin is covalently linked to suberin, and to a lesser extent, to cutin. Suberin is a major constituent of cork, and is named after the cork oak, Quercus suber. Its main function is as a barrier to movement of water and solutes.

The Casparian strip is a band-like thickening in the center of the root endodermis of vascular plants. The composition of the region is mainly suberin, lignin and some structural proteins, which are capable of reducing the diffusive apoplastic flow of water and solutes into the stele and its width varies between species. The Casparian strip is impervious to water so can control the transportation of water and inorganic salts between the cortex and the vascular bundle, preventing water and inorganic salts from being transported to the stele through the apoplast, so that it must enter the cell membrane and move to the stele through the symplastic pathway, blocking the internal and external objects of the cell. The function and function of mass transportation are similar to that of animal tissues.. The development of the Casparian strip is regulated by transcription factors such as SHORT-ROOT (SHR), SCARECROW (SCR) and MYB36, as well as polypeptide hormone synthesised by midcolumn cells.

The apoplast is the extracellular space outside of plant cell membranes, especially the fluid-filled cell walls of adjacent cells where water and dissolved material can flow and diffuse freely. Fluid and material flows occurring in any extracellular space are called apoplastic flow or apoplastic transport. The apoplastic pathway is one route by which water and solutes are transported and distributed to different places through tissues and organs, contrasting with the symplastic pathway.

Eduard Adolf Strasburger was a Polish-German professor and one of the most famous botanists of the 19th century. He discovered mitosis in plants.

Plasmodesmata are microscopic channels which traverse the cell walls of plant cells and some algal cells, enabling transport and communication between them. Plasmodesmata evolved independently in several lineages, and species that have these structures include members of the Charophyceae, Charales, Coleochaetales and Phaeophyceae, as well as all embryophytes, better known as land plants. Unlike animal cells, almost every plant cell is surrounded by a polysaccharide cell wall. Neighbouring plant cells are therefore separated by a pair of cell walls and the intervening middle lamella, forming an extracellular domain known as the apoplast. Although cell walls are permeable to small soluble proteins and other solutes, plasmodesmata enable direct, regulated, symplastic transport of substances between cells. There are two forms of plasmodesmata: primary plasmodesmata, which are formed during cell division, and secondary plasmodesmata, which can form between mature cells.

In plants, the transpiration stream is the uninterrupted stream of water and solutes which is taken up by the roots and transported via the xylem to the leaves where it evaporates into the air/apoplast-interface of the substomatal cavity. It is driven by capillary action and in some plants by root pressure. The main driving factor is the difference in water potential between the soil and the substomatal cavity caused by transpiration.
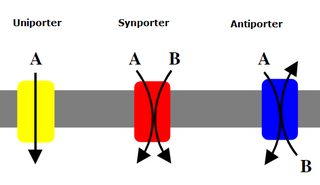
A symporter is an integral membrane protein that is involved in the transport of two different molecules across the cell membrane in the same direction. The symporter works in the plasma membrane and molecules are transported across the cell membrane at the same time, and is, therefore, a type of cotransporter. The transporter is called a symporter, because the molecules will travel in the same direction in relation to each other. This is in contrast to the antiport transporter. Typically, the ion(s) will move down the electrochemical gradient, allowing the other molecule(s) to move against the concentration gradient. The movement of the ion(s) across the membrane is facilitated diffusion, and is coupled with the active transport of the molecule(s). In symport, two molecule move in a 'similar direction' at the 'same time'. For example, the movement of glucose along with sodium ions.

A pulvinus is a joint-like thickening at the base of a plant leaf or leaflet that facilitates growth-independent movement. Pulvini are common, for example, in members of the bean family Fabaceae (Leguminosae) and the prayer plant family Marantaceae.
The pressure flow hypothesis, also known as the mass flow hypothesis, is the best-supported theory to explain the movement of sap through the phloem. It was proposed by Ernst Münch, a German plant physiologist in 1930. A high concentration of organic substances, particularly sugar, inside cells of the phloem at a source, such as a leaf, creates a diffusion gradient that draws water into the cells from the adjacent xylem. This creates turgor pressure, also known as hydrostatic pressure, in the phloem. Movement of phloem sap occurs by bulk flow from sugar sources to sugar sinks. The movement in phloem is bidirectional, whereas, in xylem cells, it is unidirectional (upward). Because of this multi-directional flow, coupled with the fact that sap cannot move with ease between adjacent sieve-tubes, it is not unusual for sap in adjacent sieve-tubes to be flowing in opposite directions.

Osmosis is the spontaneous net movement or diffusion of solvent molecules through a selectively-permeable membrane from a region of high water potential to a region of low water potential, in the direction that tends to equalize the solute concentrations on the two sides. It may also be used to describe a physical process in which any solvent moves across a selectively permeable membrane separating two solutions of different concentrations. Osmosis can be made to do work. Osmotic pressure is defined as the external pressure required to be applied so that there is no net movement of solvent across the membrane. Osmotic pressure is a colligative property, meaning that the osmotic pressure depends on the molar concentration of the solute but not on its identity.
Vacuolar pathway: Movement of water molecules in plant cells via the vacuoles located in the cytoplasm of the cell. The water molecules encounter high resistance however and as a result little flow usually occurs making this pathway insignificant, the Apoplast pathway and Symplast pathway being the major pathways for movement of water in plants. Water moves by osmosis across the vacuoles of the cells of the root system. The water moves down a concentration gradient from the soil solution to the xylem. The vacuolar pathway can also be considered a symplast pathway.

Pits are relatively thinner portions of the cell wall that adjacent cells can communicate or exchange fluid through. Pits are characteristic of cell walls with secondary layers. Generally each pit has a complementary pit opposite of it in the neighboring cell. These complementary pits are called "pit pairs".

The exodermis is a physiological barrier that has a role in root function and protection. The exodermis is a membrane of variable permeability responsible for the radial flow of water, ions, and nutrients. It is the outer layer of a plant's cortex. The exodermis serves a double function as it can protect the root from invasion by foreign pathogens and ensures that the plant does not lose too much water through diffusion through the root system and can properly replenish its stores at an appropriate rate.

Phloem loading is the process of loading carbon into the phloem for transport to different 'sinks' in a plant. Sinks include metabolism, growth, storage, and other processes or organs that need carbon solutes to persist. It can be a passive process, relying on a pressure gradient to generate diffusion of solutes through the symplast, or an active process, requiring energy to create membrane-bound transporter proteins that move solutes through the apoplast against a gradient. Passive phloem loading transports solutes freely through plasmodesma in the symplast of the minor veins of leaves. Active transport occurs apoplastically and does not use plasmodesmata. An intermediate type of loading exists that uses symplastic transport but utilizes a size-exclusion mechanism to ensure diffusion is a one-way process between the mesophyll and phloem cells. This process is referred to as polymer-trapping, in which simple solutes such as sucrose are synthesized into larger molecules such as stachyose or raffinose in intermediary cells. The larger molecules cannot diffuse back to the mesophyll but can move into the phloem's sieve cells. Therefore, the synthesis of larger compounds uses energy and is thus 'active' but this strategy does not require specialized proteins and can still move symplastically.
