Related Research Articles

Pollen is a powdery substance produced by most types of flowers of seed plants for the purpose of sexual reproduction. It consists of pollen grains, which produce male gametes. Pollen grains have a hard coat made of sporopollenin that protects the gametophytes during the process of their movement from the stamens to the pistil of flowering plants, or from the male cone to the female cone of gymnosperms. If pollen lands on a compatible pistil or female cone, it germinates, producing a pollen tube that transfers the sperm to the ovule containing the female gametophyte. Individual pollen grains are small enough to require magnification to see detail. The study of pollen is called palynology and is highly useful in paleoecology, paleontology, archaeology, and forensics. Pollen in plants is used for transferring haploid male genetic material from the anther of a single flower to the stigma of another in cross-pollination. In a case of self-pollination, this process takes place from the anther of a flower to the stigma of the same flower.
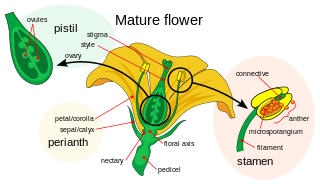
Petals are modified leaves that surround the reproductive parts of flowers. They are often brightly colored or unusually shaped to attract pollinators. All of the petals of a flower are collectively known as the corolla. Petals are usually accompanied by another set of modified leaves called sepals, that collectively form the calyx and lie just beneath the corolla. The calyx and the corolla together make up the perianth, the non-reproductive portion of a flower. When the petals and sepals of a flower are difficult to distinguish, they are collectively called tepals. Examples of plants in which the term tepal is appropriate include genera such as Aloe and Tulipa. Conversely, genera such as Rosa and Phaseolus have well-distinguished sepals and petals. When the undifferentiated tepals resemble petals, they are referred to as "petaloid", as in petaloid monocots, orders of monocots with brightly colored tepals. Since they include Liliales, an alternative name is lilioid monocots.

The stamen is the pollen-producing reproductive organ of a flower. Collectively the stamens form the androecium.

Pollination is the transfer of pollen from an anther of a plant to the stigma of a plant, later enabling fertilisation and the production of seeds, most often by an animal or by wind. Pollinating agents can be animals such as insects, for example beetles or butterflies; birds, and bats; water; wind; and even plants themselves. Pollinating animals travel from plant to plant carrying pollen on their bodies in a vital interaction that allows the transfer of genetic material critical to the reproductive system of most flowering plants. When self-pollination occurs within a closed flower. Pollination often occurs within a species. When pollination occurs between species, it can produce hybrid offspring in nature and in plant breeding work.

Pollination of fruit trees is required to produce seeds with surrounding fruit. It is the process of moving pollen from the anther to the stigma, either in the same flower or in another flower. Some tree species, including many fruit trees, do not produce fruit from self-pollination, so pollinizer trees are planted in orchards.

A pollen tube is a tubular structure produced by the male gametophyte of seed plants when it germinates. Pollen tube elongation is an integral stage in the plant life cycle. The pollen tube acts as a conduit to transport the male gamete cells from the pollen grain—either from the stigma to the ovules at the base of the pistil or directly through ovule tissue in some gymnosperms. In maize, this single cell can grow longer than 12 inches (30 cm) to traverse the length of the pistil.

Self-pollination is a form of pollination in which pollen from the same plant arrives at the stigma of a flower or at the ovule. There are two types of self-pollination: in autogamy, pollen is transferred to the stigma of the same flower; in geitonogamy, pollen is transferred from the anther of one flower to the stigma of another flower on the same flowering plant, or from microsporangium to ovule within a single (monoecious) gymnosperm. Some plants have mechanisms that ensure autogamy, such as flowers that do not open (cleistogamy), or stamens that move to come into contact with the stigma. The term selfing that is often used as a synonym, is not limited to self-pollination, but also applies to other type of self-fertilization.

Plant reproductive morphology is the study of the physical form and structure of those parts of plants directly or indirectly concerned with sexual reproduction.
Self-incompatibility (SI) is a general name for several genetic mechanisms that prevent self-fertilization in sexually reproducing organisms, and thus encourage outcrossing and allogamy. It is contrasted with separation of sexes among individuals (dioecy), and their various modes of spatial (herkogamy) and temporal (dichogamy) separation.

Heterostyly is a unique form of polymorphism and herkogamy in flowers. In a heterostylous species, two or three morphological types of flowers, termed "morphs", exist in the population. On each individual plant, all flowers share the same morph. The flower morphs differ in the lengths of the pistil and stamens, and these traits are not continuous. The morph phenotype is genetically linked to genes responsible for a unique system of self-incompatibility, termed heteromorphic self-incompatibility, that is, the pollen from a flower on one morph cannot fertilize another flower of the same morph.
Geitonogamy is a type of self-pollination. Geitonogamous pollination is sometimes distinguished from the fertilizations that can result from it, geitonogamy. If a plant is self-incompatible, geitonogamy can reduce seed production.

The tribe Euglossini, in the subfamily Apinae, commonly known as orchid bees or euglossine bees, are the only group of corbiculate bees whose non-parasitic members do not all possess eusocial behavior.

A flower, also known as a bloom or blossom, is the reproductive structure found in flowering plants. Flowers consist of a combination of vegetative organs – sepals that enclose and protect the developing flower, petals that attract pollinators, and reproductive organs that produce gametophytes, which in flowering plants produce gametes. The male gametophytes, which produce sperm, are enclosed within pollen grains produced in the anthers. The female gametophytes are contained within the ovules produced in the carpels.

Pollination syndromes are suites of flower traits that have evolved in response to natural selection imposed by different pollen vectors, which can be abiotic or biotic, such as birds, bees, flies, and so forth through a process called pollinator-mediated selection. These traits include flower shape, size, colour, odour, reward type and amount, nectar composition, timing of flowering, etc. For example, tubular red flowers with copious nectar often attract birds; foul smelling flowers attract carrion flies or beetles, etc.
Plant reproduction is the production of new offspring in plants, which can be accomplished by sexual or asexual reproduction. Sexual reproduction produces offspring by the fusion of gametes, resulting in offspring genetically different from either parent. Asexual reproduction produces new individuals without the fusion of gametes, resulting in clonal plants that are genetically identical to the parent plant and each other, unless mutations occur.
Tristyly is a rare floral polymorphism that consists of three floral morphs that differ in regard to the length of the stamens and style within the flower. This type of floral mechanism is thought to encourage outcross pollen transfer and is usually associated with heteromorphic self-incompatibility to reduce inbreeding. It is an example of heterostyly and reciprocal herkogamy, like distyly, which is the more common form of heterostyly. Darwin first described tristylous species in 1877 in terms of the incompatibility of these three morphs.
This page provides a glossary of plant morphology. Botanists and other biologists who study plant morphology use a number of different terms to classify and identify plant organs and parts that can be observed using no more than a handheld magnifying lens. This page provides help in understanding the numerous other pages describing plants by their various taxa. The accompanying page—Plant morphology—provides an overview of the science of the external form of plants. There is also an alphabetical list: Glossary of botanical terms. In contrast, this page deals with botanical terms in a systematic manner, with some illustrations, and organized by plant anatomy and function in plant physiology.

The monocots are one of the two major groups of flowering plants, the other being the dicots. In order to reproduce they utilize various strategies such as employing forms of asexual reproduction, restricting which individuals they are sexually compatible with, or influencing how they are pollinated. Nearly all reproductive strategies that evolved in the dicots have independently evolved in monocots as well. Despite these similarities and their close relatedness, monocots and dicots have distinct traits in their reproductive biologies.

A mixed mating system, also known as “variable inbreeding” a characteristic of many hermaphroditic seed plants, where more than one means of mating is used. Mixed mating usually refers to the production of a mixture of self-fertilized (selfed) and outbred (outcrossed) seeds. Plant mating systems influence the distribution of genetic variation within and among populations, by affecting the propensity of individuals to self-fertilize or cross-fertilize . Mixed mating systems are generally characterized by the frequency of selfing vs. outcrossing, but may include the production of asexual seeds through agamospermy. The trade offs for each strategy depend on ecological conditions, pollinator abundance and herbivory and parasite load. Mating systems are not permanent within species; they can vary with environmental factors, and through domestication when plants are bred for commercial agriculture.

The pollination of orchids is a complex chapter in the biology of this family of plants that are distinguished by the complexity of their flowers and by intricate ecological interactions with their pollinator agents. It has captured the attention of numerous scientists over time, including Charles Darwin, father of the theory of evolution by natural selection. Darwin published in 1862 the first observations of the fundamental role of insects in orchid pollination, in his book The Fertilization of Orchids. Darwin stated that the varied stratagems orchids use to attract their pollinators transcend the imagination of any human being.
References
- ↑ Biology textbook for XII. Nation Council of Educational Research and Training. 2006. p. 28. ISBN 81-7450-639-X.
- ↑ Darwin, Charles (August 2006). More Letters of Charles Darwin, Volume 2. Echo Library. p. 668. ISBN 978-1-4068-0482-9 . Retrieved 25 February 2012.