
Brown algae, comprising the class Phaeophyceae, are a large group of multicellular algae, including many seaweeds located in colder waters within the Northern Hemisphere. Brown algae are the major seaweeds of the temperate and polar regions. They are dominant on rocky shores throughout cooler areas of the world. Most brown algae live in marine environments, where they play an important role both as food and as a potential habitat. For instance, Macrocystis, a kelp of the order Laminariales, may reach 60 m (200 ft) in length and forms prominent underwater kelp forests. Kelp forests like these contain a high level of biodiversity. Another example is Sargassum, which creates unique floating mats of seaweed in the tropical waters of the Sargasso Sea that serve as the habitats for many species. Many brown algae, such as members of the order Fucales, commonly grow along rocky seashores. Some members of the class, such as kelps, are used by humans as food.

Fucus is a genus of brown algae found in the intertidal zones of rocky seashores almost throughout the world.

Coralline algae are red algae in the order Corallinales. They are characterized by a thallus that is hard because of calcareous deposits contained within the cell walls. The colors of these algae are most typically pink, or some other shade of red, but some species can be purple, yellow, blue, white, or gray-green. Coralline algae play an important role in the ecology of coral reefs. Sea urchins, parrot fish, and limpets and chitons feed on coralline algae. In the temperate Mediterranean Sea, coralline algae are the main builders of a typical algal reef, the Coralligène ("coralligenous"). Many are typically encrusting and rock-like, found in marine waters all over the world. Only one species lives in freshwater. Unattached specimens may form relatively smooth compact balls to warty or fruticose thalli.

Atractophora hypnoides is a rare red alga (Rhodophyta) found in the British Isles, France and some Atlantic Islands and is the only species of the genus found in the British Isles. It is attached to the rock or other algae by a small basal disc and is much branched with downgrowing filaments which enclose the main branch or axis forming a cortex. Short filaments of limited growth radiate in whorls from the axis and frequently convert into hairs. The spreading filaments grow irregularly in a diffuse manner. Microscope examination is required for identification.

Polysiphonia is a genus of filamentous red algae with about 19 species on the coasts of the British Isles and about 200 species worldwide, including Crete in Greece, Antarctica and Greenland. Its members are known by a number of common names. It is in the order Ceramiales and family Rhodomelaceae.

Conceptacles are specialized cavities of marine and freshwater algae that contain the reproductive organs. They are situated in the receptacle and open by a small ostiole. Conceptacles are present in Corallinaceae, and Hildenbrandiales, as well as the brown Fucales. In the Fucales there is no haploid phase in the reproductive cycle and therefore no alternation of generations. The thallus is a sporophyte. The diploid plants produce male (antheridia) and female (oogonia) gametangia by meiosis. The gametes are released into the surrounding water; after fusion, the zygote settles and begins growth.

Calliarthron is a genus containing two species of thalloid intertidal alga. Specimens can reach around 30 cm in size. The thalli take a crustose form. The organisms lack secondary pit connections. Calliarthron reproduces by means of conceptacles; it produces tetraspores, dispores and carpospores. The genus has lignin and contains secondary cell walls, traits which are normally associated with the vascular plants. It is similar to the genus Bossiella.
Bangia is an extant genus of division Rhodophyta that grows in marine or freshwater habitats. Bangia has small thalli with rapid growth and high reproductive output, and exhibits behavior characteristic of r-selected species. The plants are attached by down-growing rhizoids, usually in dense purple-black to rust-colored clumps. The chloroplasts of Bangia, like others in the division Rhodophyta, contain chlorophyll a and sometimes chlorophyll d, as well as accessory pigments such as phycobilin pigments and xanthophylls. Depending on the relative proportions of these pigments and the light conditions, the overall color of the plant can range from green to red to purple to grey; however, the red pigment, phycoerythrin, is usually dominant.
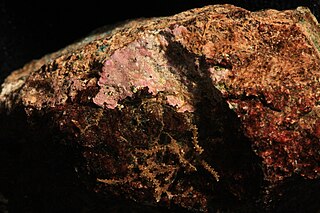
Hildenbrandia is a genus of thalloid red alga comprising about 26 species. The slow-growing, non-mineralized thalli take a crustose form. Hildenbrandia reproduces by means of conceptacles and produces tetraspores.
The epithallium or epithallus is the outer layer of a crustose coralline alga, which in some species is periodically shed to prevent organisms from attaching to and overgrowing the alga.

Amphiroa is a genus of thalloid red algae under the family Corallinaceae.

Jania is a genus of red macroalgae with hard, calcareous, branching skeletons in the family Corallinaceae.

Spongites yendoi is a species of crustose red seaweed with a hard, calcareous skeleton in the family Corallinaceae. It is found on the lower shore as part of a diverse community in the southeastern Atlantic Ocean and the Indo-Pacific Ocean.

Polysiphonia lanosa is a common species of the red algae (Rhodophyta) often to be found growing on Ascophyllum nodosum.

Notheia anomala is a macroalga in the family Notheiceae and the brown algae order Fucales. It is an obligate epiphyte of another brown algae, Hormosira banksii.

Phycodrys rubens is a red marine alga of up to 30 cm long.

Lithophyllum orbiculatum is a species of thalloid coralline algae, which are a red algae whose cell walls contain calcareous deposits.

Callithamniaceae is a family of red algae (Rhodophyta) in the order Ceramiales. The family was first described by Friedrich Traugott Kützing in 1843.

Hildenbrandia rubra is a marine species of thalloid red alga. It forms thin reddish crusts on rocks and pebbles in the intertidal zone and the shallow subtidal zone. It is a common species with a cosmopolitan distribution, and is able to tolerate a wide range of conditions.
Crustaphytum is a genus of red alga first discovered in Taoyuan algal reefs by Taiwanese scientists. The epithet “crusta” refers to crustose thallus and “phytum” refers to plant. Belonging to the family Hapalidiaceae in the order Hapalidiales, Crustaphytum is one kind of crustose coralline algae.




