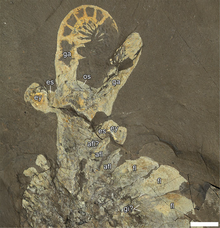
Like other radiodonts, Anomalocaris had swimming flaps running along its body, large compound eyes, and a single pair of segmented, frontal appendages, which in Anomalocaris were used to grasp prey. Measuring up to 38cm (1.25ft) long excluding frontal appendages and tail fan,[4]A. canadensis is one of the largest animals of the Cambrian, and thought to be one of the earliest examples of an apex predator,[5][6] though others have been found in older Cambrian lagerstätten deposits.
Since the original description in late 19th century,[7] the frontal appendages were the only known fossilized parts and misidentified as the body parts of other animals.[8] Its radiodont affinity was revealed in 1980s, specifically in a 1985 journal article by Harry B. Whittington and Derek Briggs.[9] The trunk and mouth were reconstructed after another radiodont genus until the corrections done in 1996[8] and 2012.[10] It is the type genus of Anomalocarididae, a family which previously included all radiodonts but recently only Anomalocaris and a few closely-related taxa.[3]
Discovery and identification
Frontal appendage of Anomalocaris canadensis (top) and mouthpiece of Peytoia nathorsti (bottom) from British Columbia. The latter was originally assigned to the former species.
From the start, Anomalocaris fossil was misidentified, followed by a series of misidentifications and taxonomic revisions.[9][8] As Stephen Jay Gould, who popularised the Cambrian explosion in his 1989 book Wonderful Life, appropriately described:
[The story of Anomalocaris is] a tale of humor, error, struggle, frustration, and more error, culminating in an extraordinary resolution that brought together bits and pieces of three "phyla" in a single reconstructed creature, the largest and fiercest of Cambrian organisms.[11]
Anomalocaris fossils were first collected in 1886[8] by Richard G. McConnell of the Geological Survey of Canada (GSC). Having been informed of rich fossils at the Stephen Formation in British Columbia, McConnell climbed Mount Stephen on 13 September 1886.[12][13] He found abundant trilobites, along with two unknown specimens.[7] In August 1891, Henri-Marc Ami, Assistant Palaeontologist at GSC, collected many trilobites and brachiopod fossils,[14] along with 48 more of the unknown specimens.[15] The fifty specimens were examined and described in 1892 by GSC paleontologist Joseph Frederick Whiteaves.[16][17] Whiteaves interpreted them as the abdomens of phyllocaridcrustaceans, and gave the full scientific name Anomalocaris canadensis. He describes the crustacean characters:
Body or abdominal segments, which, in all the specimens collected, are abnormally flattened laterally, a little higher or deeper than long, broader above than below, the pair of ventral appendages proceeding from each, nearly equal in height or depth to the segment itself... The generic name Anomalocaris (from ανώμαλος, unlike,—καρίς, a shrimp, i.e., unlike other other shrimps) [the species name referring to Canada] is suggested by the unusual shape of the uropods or ventral appendages of the body segments and the relative position of the caudal spine.[7]
In 1928, Danish paleontologist Kai Henriksen proposed that Tuzoia, a Burgess Shale arthropod which was known only from the carapace, represented the missing front half of Anomalocaris.[8] The artists Elie Cheverlange and Charles R. Knight followed this interpretation in their depictions of Anomalocaris.[8]
Not known to scientists at the time, the body parts of relatives of Anomalocaris had already been described but not recognized as such. The first fossilized mouth of such a kind of animal was discovered by Charles Doolittle Walcott, who mistook it for a jellyfish and placed it in the genus Peytoia. Walcott also discovered a frontal appendage but failed to realize the similarities to Whiteaves' discovery and instead identified it as feeding appendage or tail of the coexisted Sidneyia.[18] In the same publication in which he named Peytoia, Walcott named Laggania, a taxon that he interpreted as a holothurian.
In 1966, the Geological Survey of Canada began a comprehensive revision of the Burgess Shale fossil record, led by Cambridge University paleontologist Harry B. Whittington.[8] In the process of this revision, Whittington and his students Simon Conway Morris and Derek Briggs would discover the true nature of Anomalocaris and its relatives, but not without contributing to the history of misinterpretations first.[18] In 1978, Conway Morris recognized that the mouthparts of Laggania were identical to Peytoia, but concluded that Laggania was a composite fossil made up of Peytoia and the sponge Corralio undulata.[19] In 1979, Briggs recognized that the fossils of Anomalocaris were appendages, not abdomens, and proposed that they were the walking legs of a giant arthropod, and that the feeding appendage Walcott had assigned to Sidneyia was the feeding appendage of similar animal, referred to as "appendage F".[20] Later, while clearing what he thought was an unrelated specimen, Harry B. Whittington removed a layer of covering stone to discover the unequivocally connected frontal appendage identical to Anomalocaris and mouthpart similar to Peytoia.[18][21] Whittington linked the two species, but it took several more years for researchers to realize that the continuously juxtaposed Peytoia, Laggania and frontal appendages (Anomalocaris and "appendage F") actually represented a single group of enormous creatures.[9] The two genera have now been placed into the order Radiodonta[8] and are commonly known as radiodonts or anomalocaridids. Since Peytoia was named first, it is the accepted correct name for the entire animal. However, the original frontal appendage was from a larger species distinct from Peytoia and "Laggania" and therefore retains the name Anomalocaris.[10]
In 2011[22] and 2020,[23]compound eyes of Anomalocaris were recovered from a paleontological dig at Emu Bay on Kangaroo Island, Australia, proving that Anomalocaris was indeed an arthropod as had been suspected. The find also indicated that advanced arthropod eyes had evolved very early, before the evolution of jointed legs or hardened exoskeletons.[22]
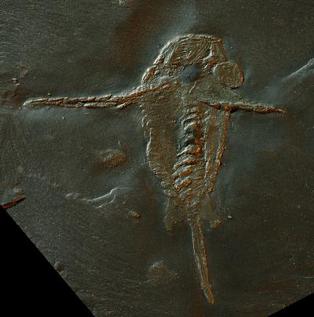
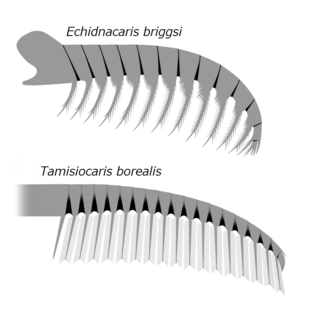
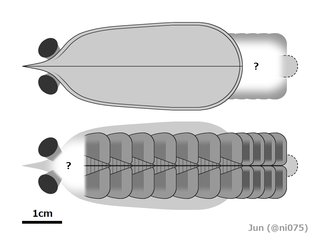
In 2021, "A." saron[24] and "A." magnabasis[25] were reassigned to the new genus Houcaris, in the family Tamisiocarididae.[26] In the same year, "A." pennsylvanica was reassigned to the genus Lenisicaris.[3] In 2022, specimen ELRC 20001 that was treated as an unnamed species of Anomalocaris or whole-body specimen of A. saron got a new genus, Innovatiocaris.[27] In 2023, "A". kunmingensis was reassigned to the new genus Guanshancaris in the family Amplectobeluidae.[28] Multiple phylogenetic analyses also suggested that "A". briggsi (tamisiocaridid) was not a species of Anomalocaris either,[29][30][31][4][32][33][34] and it was reassigned to the genus Echidnacaris in the family Tamisiocarididae in 2023.[2] In the same 2023 study, a new species of Anomalocaris, A. daleyae, was described based on remains found in the Emu Bay Shale in Australia.[2]
Description
Size estimation of Anomalocaris.
For the time in which it lived, Anomalocaris was gigantic. A. canadensis is estimated to be up to 37.8cm (1.24ft) long excluding frontal appendages and tail fans.[4][27] Previous estimation up to 1m (3.3ft)[20] is unlikely based on the ratio of body parts (body length measured only about 2 times the length of frontal appendage in A. canadensis, respectively[4]) and the size of largest frontal appendage (up to 18 centimetres (7.1 inches) in length when extended).[18][4]
Anomalocaris propelled itself through the water by undulating the flexible flaps on the sides of its body.[35] Each flap sloped below the one more posterior to it,[36] and this overlapping allowed the lobes on each side of the body to act as a single "fin", maximizing the swimming efficiency.[35] The construction of a remote-controlled model showed this mode of swimming to be intrinsically stable,[37] implying that Anomalocaris would not have needed a complex brain to manage balance while swimming. The body was widest between the third and fifth lobe and narrowed towards the tail, with additional 3 pairs of small flaps on the constricted neck region.[8][38] It is difficult to distinguish lobes near the tail, making an accurate count difficult.[36] For the main trunk flaps, the type species A. canadensis had 13 pairs.[38]
Oral cones of Anomalocaris (A), Peytoia (B) and Hurdia (C), the former showing unique triradial, tuberculated and wrinkled structures.
The mobility of radiodont trunk appendages (like Anomalocaris).
Anomalocaris had an unusual disk-like mouth known as oral cone. The oral cone was composed of several plates organized triradially. Three of the plates were quite large. Three to four medium sized plates could be found between each of the large plates, and several small plates between them. Most of the plates wrinkled and have scale-like tubercles near the mouth opening.[10][39] Such an oral cone is very different from those of a typical hurdiid radiodont like Peytoia and Hurdia, which is smooth and tetraradial.[10][33] As a shared character across radiodonts, Anomalocaris also had three sclerites on the top and side of its head.[33] The top one, known as a head shield, dorsal carapace or H-element, was shaped like an laterally-elongated[40] oval, with a distinct rim on the outer edge.[38] The remaining two lateral sclerites, known as P-elements, were also ovoid, but connected by a bar-like outgrowth.[33] The P-elements were previously misinterpreted as two huge compound eyes.[38][33]
Frontal appendages of Anomalocaris, with examples from multiple species.
Two large frontal appendage were positioned in front of the mouth, at the front of the head.[8] Each frontal appendage of Anomalocaris usually had 14 podomeres (segmental units, at least 1 for shaft and 13 for distal articulated region), with each appendage being laterally-flattened (taller than wide).[38] Most podomeres were tipped with a pair of endites (ventral spines).[38] The endites themselves were both equipped with multiple auxiliary spines, which branches off from the anterior and posterior margin of the endites.[24][39][41][38][1]
The tail was a large tail fan, composed of three[8][38] pairs of large, lateral fin-shaped lobes and one terminal lobe-like tailpiece.[38] Previous studies suggest the tail fan was used to propel it through Cambrian waters,[21][35] while further hydrodynamic study rather suggest it was more adapted to provide steering function.[42] The gills of the animal, in the form of long, thin, hair-like structures known as lanceolate blades, were arranged in rows forming setal blades. The setal blades were attached by their margin to the top side of the animal, two setal blades per body segment. A divide ran down the middle, separating the gills.[38]
Based on fossilized eyes from the Emu Bay Shale, which belong to the species Anomalocarisdaleyae,[2] the stalked eyes of Anomalocaris were 30 times more powerful than those of trilobites, long thought to have had the most advanced eyes of any contemporary species. With one specimen having over 24,000 lenses in one eye, the resolution of the 3-centimetre-wide (1.2in) eyes would have been rivalled only by that of the modern dragonfly, which has 28,000 lenses in each eye.[22][23] Additionally, estimation of ecdysozoanopsins suggest that Anomalocaris may have had dichromaticcolor vision.[43]
Paleobiology
Diet
Grasping movement of the frontal appendage of A. canadensis.
The interpretation of Anomalocaris as an active predator is widely accepted throughout the history of research,[9][8][10] as its raptorial frontal appendages and mid-gut glands strongly suggest a predatory lifestyle.[44][45][5] In the case of A. canadensis, its outstanding size amongst Burgess Shale fauna also make it one of the first apex predators known to exist.[5]
However, the long-standing idea that Anomalocaris fed on hard-bodied animals, especially its ability to penetrate mineralized exoskeleton of trilobites, has been questioned, with many recent studies considering it more likely that Anomalocaris exclusively hunted soft-bodied prey.[46][10][5][6] Some Cambrian trilobites have been found with round or W-shaped "bite" marks, which were identified as being the same shape as the mouthparts of Peytoia (previously misidentified as those of Anomalocaris[47][10]). Stronger evidence that Anomalocaris ate trilobites comes from coprolite, which contain trilobite parts and are so large that the radiodonts are the only known organism from that period large enough to have produced them.[47] However, since Anomalocaris lacks any mineralized tissue, it seemed unlikely that it would be able to penetrate the hard, calcified exoskeleton of trilobites.[47] Rather, the coprolites may have been produced by different organisms, such as the trilobites of the genus Redlichia.[39] Another suggested possibility was that Anomalocaris fed by grabbing one end of their prey in its oral cone while using its frontal appendages to quickly rock the other end of the animal back and forth. This produced stresses that exploited the weaknesses of arthropod cuticles, causing the prey's exoskeleton to rupture and allowing the predator to access its innards.[47] This behaviour was originally thought to have provided an evolutionary pressure for trilobites to roll up, to avoid being flexed until they snapped.[47]
Ecological reconstruction of Anomalocaris hunting Isoxys, after posture estimated in Bicknell et al. (2023)
The lack of wear on radiodont mouthparts suggests they did not come into regular contact with mineralized trilobite shells, and were possibly better suited to feeding on smaller, soft-bodied organisms by suction, since they would have experienced structural failure if they were used against the armour of trilobites.[46][39]A. canadensis was suggested to have been capable of feeding on organisms with hard exoskeletons due to the short, robust spines on its frontal appendages.[39][25] However, this conclusion is solely based on the comparison with the fragile frontal appendages of suspension feeding radiodonts (e.g. Echidnacaris and Houcaris spp.).[26] The typical lack of damage to the endites on the frontal appendages of A. canadensis (with damage only present on a single specimen) suggests that they were not used to grasp hard-shelled prey.[6] As opposed to Peytoia whose oral cone is more rectangular with short protruding spines, the oral cone of A. canadensis has a smaller and more irregular opening, not permitting strong biting motions, and indicating a suction-feeding behavior to suck in softer organisms.[10] Three-dimensional modelling of various radiodont frontal appendages also suggest that A. canadensis is more capable to prey on smaller (2–5cm in diameter), active, soft-bodied animals (e.g. vetulicolian; free-swimming arthropods like isoxyids and hymenocarines; Nectocaris).[5][6]
Bicknell et al. (2023) examined the frontal appendages of Anomalocaris, suggesting it was an active nektonic apex predator. Postured with the frontal appendages outstretched, Anomalocaris would have been able to swim with maximized speed, similar to modern predatory water bugs. Its eyes would be suitable to hunt prey in well-lit waters. Anomalocaris would have hunted various free-swimming animals since there are a large diversity of nektonic and pelagic soft-bodied animals. It probably would have not hunted benthic animals like trilobites, considering the possibility of damaging the frontal appendages on the substrate while trying to grab prey from seafloor at speed. Instead, other animals such as other radiodonts (e.g. Hurdia, Cambroraster, Titanokorys, Stanleycaris) and artiopods (e.g. Sidneyia) would have been benthic predators in the Burgess Shale.[5][6]
Anomalocaris canadensis lived in the Burgess Shale in relatively great numbers.[1] In the Burgess Shale, Anomalocaris is more common in the older sections, notably the Mount Stephen trilobite beds. However, in the younger sections, such as the Phyllopod bed, Anomalocaris could reach much greater sizes; roughly twice the size of its older, trilobite bed relatives. These rare giant specimens have previously been referred to a separate species, Anomalocaris gigantea; however, the validity of this species has been called into question,[20] and is currently synonymized to A. canadensis.[38]
Other unnamed species of Anomalocaris live in vastly different environments.[3] For example, Anomalocaris cf. canadensis (JS-1880) lived in the Maotianshan Shales,[3] a shallow tropical sea or even being delta[48] in what is now modern China. Anomalocaris daleyae (Emu Bay Shale) lived in a comparable environment; the shallow, tropical waters of Cambrian Australia.[3] The Maotianshan Shale and the Emu Bay Shale are very close in proximity, being separated by a small landmass, far from the Burgess Shale.[3] These two locations also included "Anomalocaris" kunmingensis and "Anomalocaris" briggsi respectively, species that previously attributed[49][50][39][51] but taxonomically unlikely to be a member of Anomalocaris nor even Anomalocarididae.[3][52]
1 2 3 4 5 6 7 8 9 10 Wu Y, Ma J, Lin W, Sun A, Zhang X, Fu D (2021). "New anomalocaridids (Panarthropoda: Radiodonta) from the lower Cambrian Chengjiang Lagerstätte: Biostratigraphic and paleobiogeographic implications". Palaeogeography, Palaeoclimatology, Palaeoecology. 569: Article 110333. Bibcode:2021PPP...569k0333W. doi:10.1016/j.palaeo.2021.110333. S2CID233565727.
↑ Whiteaves JF (1892). "Description of a new genus and species of phyllocarid Crustacea from the Middle Cambrian of Mount Stephen, B. C.". The Canadian Record of Science. 5 (4).
↑ Conway Morris S (1978). "Laggania cambria Walcott: A Composite Fossil". Journal of Paleontology. 52 (1): 126–131. JSTOR1303799.
1 2 3 Briggs DE (1979). "Anomalocaris, the largest known Cambrian arthropod". Palaeontology. 22 (3): 631–664.
1 2 Conway Morris S (1998). The crucible of creation: the Burgess Shale and the rise of animals. Oxford [Oxfordshire]: Oxford University Press. pp.56–9. ISBN978-0-19-850256-2.
1 2 Xian-Guang H, Bergström J, Ahlberg P (1995-09-01). "Anomalocaris and other large animals in the lower Cambrian Chengjiang fauna of southwest China". GFF. 117 (3): 163–183. doi:10.1080/11035899509546213. ISSN1103-5897.
1 2 Wu Y, Fu D, Ma J, Lin W, Sun A, Zhang X (2021). "Houcaris gen. nov. from the early Cambrian (Stage 3) Chengjiang Lagerstätte expanded the palaeogeographical distribution of tamisiocaridids (Panarthropoda: Radiodonta)". PalZ. 95 (2): 209–221. doi:10.1007/s12542-020-00545-4. ISSN1867-6812. S2CID235221043.
1 2 Zeng H, Zhao F, Zhu M (2022-09-07). "Innovatiocaris, a complete radiodont from the early Cambrian Chengjiang Lagerstätte and its implications for the phylogeny of Radiodonta". Journal of the Geological Society. 180. doi:10.1144/jgs2021-164. ISSN0016-7649. S2CID252147346.
↑ Liu Q (2013-09-01). "The first discovery of anomalocaridid appendages from the Balang Formation (Cambrian Series 2) in Hunan, China". Alcheringa: An Australasian Journal of Palaeontology. 37 (3): 338–343. doi:10.1080/03115518.2013.753767. ISSN0311-5518. S2CID129212098.
↑ Sheppard KA, Rival DE, Caron JB (October 2018). "On the Hydrodynamics of Anomalocaris Tail Fins". Integrative and Comparative Biology. 58 (4): 703–711. doi:10.1093/icb/icy014. hdl:1974/22737. PMID29697774.
Wikimedia Commons has media related to Anomalocaris.
"Anomalocaris canadensis". Burgess Shale Fossil Gallery. Virtual Museum of Canada. 2011. Archived from the original on 2020-11-12. Retrieved 2023-01-21.
Opabinia regalis is an extinct, stem group arthropod found in the Middle Cambrian Burgess Shale Lagerstätte of British Columbia. Opabinia was a soft-bodied animal, measuring up to 7 cm in body length, and its segmented trunk had flaps along the sides and a fan-shaped tail. The head shows unusual features: five eyes, a mouth under the head and facing backwards, and a clawed proboscis that probably passed food to the mouth. Opabinia probably lived on the seafloor, using the proboscis to seek out small, soft food. Fewer than twenty good specimens have been described; 3 specimens of Opabinia are known from the Greater Phyllopod bed, where they constitute less than 0.1% of the community.
Dinocaridida is a proposed fossil taxon of basal arthropods that flourished in the Cambrian period with occasional Ordovician and Devonian records. Characterized by a pair of frontal appendages and series of body flaps, the name of Dinocaridids refers to the suggested role of some of these members as the largest marine predators of their time. Dinocaridids are occasionally referred to as the 'AOPK group' by some literatures, as the group compose of Radiodonta, Opabiniidae, and the "gilled lobopodians" Pambdelurion and Kerygmachelidae. It is most likely paraphyletic, with Kerygmachelidae and Pambdelurion more basal than the clade compose of Opabiniidae, Radiodonta and other arthropods.
The Emu Bay Shale is a geological formation in Emu Bay, South Australia, containing a major Konservat-Lagerstätte. It is one of two in the world containing Redlichiidan trilobites. The Emu Bay Shale is dated as Cambrian Series 2, Stage 4, correlated with the upper Botomian Stage of the Lower Cambrian.
Peytoia is a genus of hurdiid radiodont, an early diverging order of stem-group arthropods, that lived in the Cambrian period, containing two species, Peytoia nathorsti from the Miaolingian of Canada and Peytoia infercambriensis from Poland, dating to Cambrian Stage 3. Its two frontal appendages had long bristle-like spines, it had no fan tail, and its short stalked eyes were behind its large head.
Amplectobelua is an extinct genus of late Early Cambrian amplectobeluid radiodont, a group of stem arthropods that mostly lived as free-swimming predators during the first half of the Paleozoic Era.
Anomalocarididae is an extinct family of Cambrian radiodonts, a group of stem-group arthropods.
Parapeytoia is a genus of Cambrian arthropod. The type and only described species is Parapeytoia yunnanensis, lived over 518 million years ago in the Maotianshan shales of Yunnan, China. Unidentified fossils from the same genus also had been discovered from the nearby Wulongqing Formation.
Peytoia infercambriensis is a species of hurdiid radiodont in the genus Peytoia.
A number of assemblages bear fossil assemblages similar in character to that of the Burgess Shale. While many are also preserved in a similar fashion to the Burgess Shale, the term "Burgess Shale-type fauna" covers assemblages based on taxonomic criteria only.
Schinderhannes bartelsi is a species of hurdiid radiodont (anomalocaridid) known from one specimen from the lower Devonian Hunsrück Slates. Its discovery was astonishing because previously, radiodonts were known only from exceptionally well-preserved fossil beds (Lagerstätten) from the Cambrian, 100 million years earlier.
Radiodonta is an extinct order of stem-group arthropods that was successful worldwide during the Cambrian period. They may be referred to as radiodonts, radiodontans, radiodontids, anomalocarids, or anomalocaridids, although the last two originally refer to the family Anomalocarididae, which previously included all species of this order but is now restricted to only a few species. Radiodonts are distinguished by their distinctive frontal appendages, which are morphologically diverse and used for a variety of functions. Radiodonts included the earliest large predators known, but they also included sediment sifters and filter feeders. Some of the most famous species of radiodonts are the Cambrian taxa Anomalocaris canadensis, Hurdia victoria, Peytoia nathorsti, Titanokorys gainessii, Cambroraster falcatus and Amplectobelua symbrachiata, the Ordovician Aegirocassis benmoulai and the Devonian Schinderhannes bartelsi.
Hurdia is an extinct genus of hurdiid radiodont that lived 505 million years ago during the Cambrian Period. Fossils have been found in North America, China and the Czech Republic.
Cucumericrus ("cucumber-leg") is an extinct genus of stem-arthropod. The type and only species is Cucumericrus decoratus, with fossils discovered from the Maotianshan Shales of Yunnan, China.
Tamisiocarididae is a family of radiodonts, extinct marine animals related to arthropods, that bore finely-spined appendages that were presumably used in filter-feeding. When first discovered, the clade was named Cetiocaridae after a speculative evolution artwork, Bearded Ceticaris by John Meszaros, that depicted a hypothetical filter-feeding radiodont at a time before any were known to exist. However, the family name was not valid according to the International Code of Zoological Nomenclature, as no real genus named "Cetiocaris" exists, and in 2019 it was formally replaced by the name Tamisiocarididae, after the only valid genus of the clade at the time. The family is only known from Series 2 of the Cambrian, unlike other radiodont families, which persisted longer into the Cambrian. All known species would have lived in tropical or subtropical waters, suggesting a preference for warmer waters.
Amplectobeluidae is a clade of Cambrian radiodonts. It currently includes five definitive genera, Amplectobelua, Lyrarapax, Ramskoeldia, Guanshancaris and a currently unnamed genus from the lower Cambrian aged Sirius Passet site in Greenland. There is also a potential fifth genus, Houcaris, but that genus has become problematic in terms of its taxonomic placement.
Hurdiidae is an extinct cosmopolitan family of radiodonts, a group of stem-group arthropods, which lived during the Paleozoic Era. It is the most long-lived radiodont clade, lasting from the Cambrian period to the Devonian period.
Houcaris is a possibly paraphyletic radiodont genus, tentatively assigned to either Amplectobeluidae, Anomalocarididae or Tamisiocarididae, known from Cambrian Series 2 of China and the United States. It contains two species, Houcaris saron and Houcaris magnabasis, both of which were originally named as species of the related genus Anomalocaris. The genus Houcaris was established for the two species in 2021 and honors Hou Xianguang, who had discovered and named the type species Anomalocaris saron in 1995 along with his colleagues Jan Bergström and Per E. Ahlberg.
Erratus is an extinct genus of marine arthropod from the Cambrian of China. Its type and only species is Erratus sperare. Erratus is likely one of the most basal known arthropods, and its discovery has helped scientists understand the early evolution of arthropod trunk appendages. Some of the stem-arthropods like radiodonts did not have legs, instead they had flap like appendages that helped them swim. Erratus on the other hand had not only flaps but also a set of primitive legs. It also supported the theory that the gills of aquatic arthropods probably evolved into the wings and lungs of terrestrial arthropods later in the Paleozoic.
Laminacaris is a genus of extinct stem-group arthropods (Radiodonta) that lived during the Cambrian period. It is monotypic with a single species Laminacaris chimera, the fossil of which was described from the Chengjiang biota of China in 2018. Around the same time, two specimens that were similar or of the same species were discovered at the Kinzers Formation in Pennsylvania, USA. The first specimens from China were three frontal appendages, without the other body parts.
Innovatiocaris is a genus of radiodont of uncertain family from the early Cambrian Chengjiang Lagerstätte of Yunnan Province, China. The genus contains two named species, I. maotianshanensis, known from a nearly complete individual measuring over 14.6 cm (5.7 in) and isolated frontal appendages, and I.? multispiniformis, known from a complete frontal appendage.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.