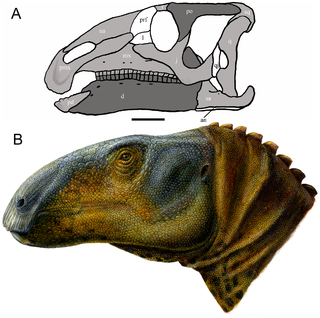
Eolambia is a genus of herbivorous hadrosauroid dinosaur from the early Late Cretaceous of the United States. It contains a single species, E. caroljonesa, named by paleontologist James Kirkland in 1998. The type specimen of Eolambia was discovered by Carole and Ramal Jones in 1993; the species name honors Carole. Since then, hundreds of bones have been discovered from both adults and juveniles, representing nearly every element of the skeleton. All of the specimens have thus far been found in Emery County, Utah, in a layer of rock known as the Mussentuchit Member of the Cedar Mountain Formation.

Huabeisaurus was a genus of dinosaur from the Late Cretaceous. It was a sauropod which lived in what is present-day northern China. The type species, Huabeisaurus allocotus, was first described by Pang Qiqing and Cheng Zhengwu in 2000. Huabeisaurus is known from numerous remains found in the 1990s, which include teeth, partial limbs and vertebrae. Due to its relative completeness, Huabeisaurus represents a significant taxon for understanding sauropod evolution in Asia. Huabeisaurus comes from Kangdailiang and Houyu, Zhaojiagou Town, Tianzhen County, Shanxi province, China. The holotype was found in the unnamed upper member of the Huiquanpu Formation, which is Late Cretaceous (?Cenomanian–?Campanian) in age based on ostracods, charophytes, and fission-track dating.

Gilmoreosaurus is the name given to a genus of dinosaur from the Cretaceous of Asia. The type species is Gilmoreosaurus mongoliensis. It is believed to be a hadrosaur or iguanodont from the Iren Dabasu Formation of Inner Mongolia, dating to 96 Ma ago. Additional specimens have been described as distinct species, including G. atavus from the Khodzhakul Formation of Uzbekistan and G. arkhangelskyi from the Bissekty Formation. However, these are based on very fragmentary remains, and their classification is dubious. An additional species, G. kysylkumense is sometimes included, though it has also been referred to the related genus Bactrosaurus.
Microhadrosaurus is a genus of hadrosaurid dinosaur from the Campanian or Maastrichtian-age Upper Cretaceous Yuanpu Formation of Guangdong, China. Although its name identifies it as a small hadrosaur, it is based on juvenile remains, and the size of the adult hadrosaur is unknown.

Nanshiungosaurus is a genus of therizinosaurid that lived in what is now Asia during the Late Cretaceous of South China. The type species, Nanshiungosaurus brevispinus, was first discovered in 1974 and described in 1979 by Dong Zhiming. It is represented by a single specimen preserving most of the cervical and dorsal vertebrae with the pelvis. A supposed and unlikely second species, "Nanshiungosaurus" bohlini, was found in 1992 and described in 1997. It is also represented by vertebrae but this species however, differs in geological age and lacks authentic characteristics compared to the type, making its affinity to the genus unsupported.

Banji is an extinct genus of oviraptorid dinosaur that lived approximately 66 million years ago during the later part of the Cretaceous Period in what is now China. It was a small, lightly built, ground-dwelling, bipedal carnivore, that was an estimated 65 cm (2.1 ft) long, as a juvenile.

Gigantoraptor is a genus of large oviraptorosaur dinosaur that lived in Asia during the Late Cretaceous period. It is known from the Iren Dabasu Formation of Inner Mongolia, where the first remains were found in 2005.

Bicentenaria is an extinct genus of carnivorous coelurosaurian theropods which lived during the early Late Cretaceous (Cenomanian) Candeleros Formation of Argentina. It contains the type species, B. argentina.

Ganzhousaurus is an extinct genus of oviraptorid dinosaur known from the Late Cretaceous Nanxiong Formation of Nankang County, Ganzhou City of Jiangxi Province, southern China. It was found in a Maastrichtian deposit and contains a single species, Ganzhousaurus nankangensis. It is distinguished by a combination of primitive and derived features.
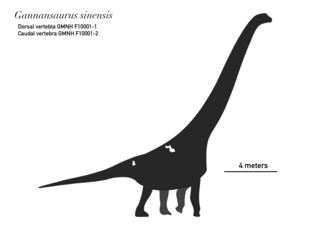
Gannansaurus is an extinct genus of somphospondylan sauropod dinosaur known from the latest Cretaceous Nanxiong Formation of Ganzhou Basin, Jiangxi Province of southern China. It is known from specimen GMNH F10001 which consists of a single, nearly complete dorsal vertebra and a mid-caudal vertebra. Gannansaurus was first named by Lü Junchang, Yi Laiping, Zhong Hui and Wei Xuefang in 2013 and the type species is Gannansaurus sinensis. Gannansaurus shares some characters with Euhelopus, indicating that it is more closely related to it rather than to other titanosauriforms.

Jiangxisaurus is an extinct genus of oviraptorid theropod dinosaur from the Late Cretaceous Nanxiong Formation of southern China. It was similar to Heyuannia, but with more strongly curved anterior claws and a thinner, frailer mandible. This find is paleontologically significant because it contributes to current knowledge about the paleogeographical distribution of oviraptorids in southern China. It was most likely an omnivorous animal along with its close relatives Nankangia and Ganzhousaurus.

Nankangia is an extinct genus of caenagnathoid oviraptorosaurian dinosaur known from the Upper Cretaceous Nanxiong Formation of Nankang County, Ganzhou City of Jiangxi Province, southeastern China. It contains a single species, Nankangia jiangxiensis. N. jiangxiensis coexisted with at least four other caenagnathoids, including but not limited to Corythoraptor, Banji, Ganzhousaurus and Jiangxisaurus. The relatively short dentary and non-downturned mandibular symphysis of Nankangia suggest that it may have been more herbivorous than carnivorous. Its diet consisted of leaves and seeds.

Qianzhousaurus is a genus of tyrannosaurid dinosaur that lived in Asia during the Maastrichtian age of the Late Cretaceous period. There is currently only one species named, the type species Qianzhousaurus sinensis, which is a member of the tribe Alioramini and most closely related to Alioramus, the only other known alioramin.
Datanglong is an extinct genus of theropod belonging to either Carcharodontosauria or Megaraptora. It existed during the Early Cretaceous (Barremian-Albian) in what is now southeastern China.

Huanansaurus is an extinct genus of oviraptorid dinosaur that lived approximately 72 million years ago, between the Campanian and Maastrichtian, during the latter part of the Cretaceous period in what is now China, in the Nanxiong Formation.

Forfexopterus is a genus of ctenochasmatid pterosaur from the Early Cretaceous Jiufotang Formation in China. It contains a single species, F. jeholensis, named from a mostly complete skeleton by Shunxing Jiang and colleagues in 2016. A second specimen, consisting of a wing, was described in 2020. While the first specimen is larger, it shows signs of being less mature than the second specimen, indicating that the developmental trajectories of Forfexopterus were variable. Like other ctenochasmatids, Forfexopterus had a long, low skull filled with many slender teeth; unlike other members of the group, however, it did not have a spatula-shaped snout tip or crests, and its teeth were more curved. A single characteristic distinguishes Forfexopterus from all other members of the wider group Archaeopterodactyloidea: of the four phalanx bones in its wing finger, the first was shorter than the second but longer than the third.

Tongtianlong is a genus of oviraptorid theropod dinosaurs that lived in the late Maastrichtian epoch of the late Cretaceous period. It contains one species, T. limosus.

Zhongjianosaurus is a genus of dromaeosaurid belonging to the Microraptoria. Believed to hail from the Yixian Formation, specifically the middle of the Jehol Biota, it is the smallest known microraptorine thus far discovered and one of the smallest non-avian theropod dinosaurs.
Mnyamawamtuka is a genus of lithostrotian titanosaur sauropod dinosaur from the Cretaceous Galula Formation in Tanzania. The type and only species is M. moyowamkia.
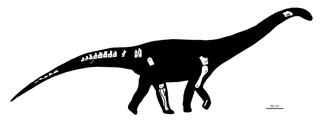
Nullotitan is a genus of lithostrotian titanosaur from the Chorrillo Formation from Santa Cruz Province in Argentina. The type and only species is Nullotitan glaciaris. It was a contemporary of the ornithopod Isasicursor which was described in the same paper.








