Related Research Articles

Within a nervous system, a neuron, neurone, or nerve cell is an electrically excitable cell that fires electric signals called action potentials across a neural network. Neurons communicate with other cells via synapses, which are specialized connections that commonly use minute amounts of chemical neurotransmitters to pass the electric signal from the presynaptic neuron to the target cell through the synaptic gap.
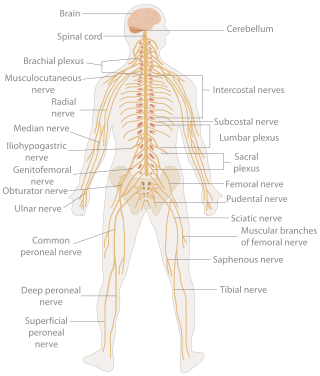
In biology, the nervous system is the highly complex part of an animal that coordinates its actions and sensory information by transmitting signals to and from different parts of its body. The nervous system detects environmental changes that impact the body, then works in tandem with the endocrine system to respond to such events. Nervous tissue first arose in wormlike organisms about 550 to 600 million years ago. In vertebrates, it consists of two main parts, the central nervous system (CNS) and the peripheral nervous system (PNS). The CNS consists of the brain and spinal cord. The PNS consists mainly of nerves, which are enclosed bundles of the long fibers, or axons, that connect the CNS to every other part of the body. Nerves that transmit signals from the brain are called motor nerves or efferent nerves, while those nerves that transmit information from the body to the CNS are called sensory nerves or afferent. Spinal nerves are mixed nerves that serve both functions. The PNS is divided into three separate subsystems, the somatic, autonomic, and enteric nervous systems. Somatic nerves mediate voluntary movement. The autonomic nervous system is further subdivided into the sympathetic and the parasympathetic nervous systems. The sympathetic nervous system is activated in cases of emergencies to mobilize energy, while the parasympathetic nervous system is activated when organisms are in a relaxed state. The enteric nervous system functions to control the gastrointestinal system. Both autonomic and enteric nervous systems function involuntarily. Nerves that exit from the cranium are called cranial nerves while those exiting from the spinal cord are called spinal nerves.
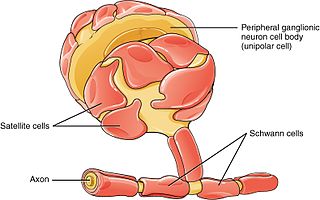
Schwann cells or neurolemmocytes are the principal glia of the peripheral nervous system (PNS). Glial cells function to support neurons and in the PNS, also include satellite cells, olfactory ensheathing cells, enteric glia and glia that reside at sensory nerve endings, such as the Pacinian corpuscle. The two types of Schwann cells are myelinating and nonmyelinating. Myelinating Schwann cells wrap around axons of motor and sensory neurons to form the myelin sheath. The Schwann cell promoter is present in the downstream region of the human dystrophin gene that gives shortened transcript that are again synthesized in a tissue-specific manner.
Erythropoietin, also known as erythropoetin, haematopoietin, or haemopoietin, is a glycoprotein cytokine secreted mainly by the kidneys in response to cellular hypoxia; it stimulates red blood cell production (erythropoiesis) in the bone marrow. Low levels of EPO are constantly secreted in sufficient quantities to compensate for normal red blood cell turnover. Common causes of cellular hypoxia resulting in elevated levels of EPO include any anemia, and hypoxemia due to chronic lung disease and mouth disease.
Nervous tissue, also called neural tissue, is the main tissue component of the nervous system. The nervous system regulates and controls body functions and activity. It consists of two parts: the central nervous system (CNS) comprising the brain and spinal cord, and the peripheral nervous system (PNS) comprising the branching peripheral nerves. It is composed of neurons, also known as nerve cells, which receive and transmit impulses, and neuroglia, also known as glial cells or glia, which assist the propagation of the nerve impulse as well as provide nutrients to the neurons.

Oligodendrocytes, also known as oligodendroglia, are a type of neuroglia whose main functions are to provide support and insulation to axons within the central nervous system (CNS) of jawed vertebrates. Their function is similar to that of Schwann cells, which perform the same task in the peripheral nervous system (PNS). Oligodendrocytes accomplish this by forming the myelin sheath around axons. Unlike Schwann cells, a single oligodendrocyte can extend its processes to cover around 50 axons, with each axon being wrapped in approximately 1 μm of myelin sheath. Furthermore, an oligodendrocyte can provide myelin segments for multiple adjacent axons.

Wallerian degeneration is an active process of degeneration that results when a nerve fiber is cut or crushed and the part of the axon distal to the injury degenerates. A related process of dying back or retrograde degeneration known as 'Wallerian-like degeneration' occurs in many neurodegenerative diseases, especially those where axonal transport is impaired such as ALS and Alzheimer's disease. Primary culture studies suggest that a failure to deliver sufficient quantities of the essential axonal protein NMNAT2 is a key initiating event.

Neurotrophins are a family of proteins that induce the survival, development, and function of neurons.

A dorsal root ganglion is a cluster of neurons in a dorsal root of a spinal nerve. The cell bodies of sensory neurons known as first-order neurons are located in the dorsal root ganglia.

Peripherin is a type III intermediate filament protein expressed mainly in neurons of the peripheral nervous system. It is also found in neurons of the central nervous system that have projections toward peripheral structures, such as spinal motor neurons. Its size, structure, and sequence/location of protein motifs is similar to other type III intermediate filament proteins such as desmin, vimentin and glial fibrillary acidic protein. Like these proteins, peripherin can self-assemble to form homopolymeric filamentous networks, but it can also heteropolymerize with neurofilaments in several neuronal types. This protein in humans is encoded by the PRPH gene. Peripherin is thought to play a role in neurite elongation during development and axonal regeneration after injury, but its exact function is unknown. It is also associated with some of the major neuropathologies that characterize amyotropic lateral sclerosis (ALS), but despite extensive research into how neurofilaments and peripherin contribute to ALS, their role in this disease is still unidentified.

Neuroprotection refers to the relative preservation of neuronal structure and/or function. In the case of an ongoing insult the relative preservation of neuronal integrity implies a reduction in the rate of neuronal loss over time, which can be expressed as a differential equation. It is a widely explored treatment option for many central nervous system (CNS) disorders including neurodegenerative diseases, stroke, traumatic brain injury, spinal cord injury, and acute management of neurotoxin consumption. Neuroprotection aims to prevent or slow disease progression and secondary injuries by halting or at least slowing the loss of neurons. Despite differences in symptoms or injuries associated with CNS disorders, many of the mechanisms behind neurodegeneration are the same. Common mechanisms of neuronal injury include decreased delivery of oxygen and glucose to the brain, energy failure, increased levels in oxidative stress, mitochondrial dysfunction, excitotoxicity, inflammatory changes, iron accumulation, and protein aggregation. Of these mechanisms, neuroprotective treatments often target oxidative stress and excitotoxicity—both of which are highly associated with CNS disorders. Not only can oxidative stress and excitotoxicity trigger neuron cell death but when combined they have synergistic effects that cause even more degradation than on their own. Thus limiting excitotoxicity and oxidative stress is a very important aspect of neuroprotection. Common neuroprotective treatments are glutamate antagonists and antioxidants, which aim to limit excitotoxicity and oxidative stress respectively.

The neuroimmune system is a system of structures and processes involving the biochemical and electrophysiological interactions between the nervous system and immune system which protect neurons from pathogens. It serves to protect neurons against disease by maintaining selectively permeable barriers, mediating neuroinflammation and wound healing in damaged neurons, and mobilizing host defenses against pathogens.

The p75 neurotrophin receptor (p75NTR) was first identified in 1973 as the low-affinity nerve growth factor receptor (LNGFR) before discovery that p75NTR bound other neurotrophins equally well as nerve growth factor. p75NTR is a neurotrophic factor receptor. Neurotrophic factor receptors bind Neurotrophins including Nerve growth factor, Neurotrophin-3, Brain-derived neurotrophic factor, and Neurotrophin-4. All neurotrophins bind to p75NTR. This also includes the immature pro-neurotrophin forms. Neurotrophic factor receptors, including p75NTR, are responsible for ensuring a proper density to target ratio of developing neurons, refining broader maps in development into precise connections. p75NTR is involved in pathways that promote neuronal survival and neuronal death.

Galanin is a neuropeptide encoded by the GAL gene, that is widely expressed in the brain, spinal cord, and gut of humans as well as other mammals. Galanin signaling occurs through three G protein-coupled receptors.

Satellite glial cells, formerly called amphicytes, are glial cells that cover the surface of neuron cell bodies in ganglia of the peripheral nervous system. Thus, they are found in sensory, sympathetic, and parasympathetic ganglia. Both satellite glial cells (SGCs) and Schwann cells are derived from the neural crest of the embryo during development. SGCs have been found to play a variety of roles, including control over the microenvironment of sympathetic ganglia. They are thought to have a similar role to astrocytes in the central nervous system (CNS). They supply nutrients to the surrounding neurons and also have some structural function. Satellite cells also act as protective, cushioning cells. Additionally, they express a variety of receptors that allow for a range of interactions with neuroactive chemicals. Many of these receptors and other ion channels have recently been implicated in health issues including chronic pain and herpes simplex. There is much more to be learned about these cells, and research surrounding additional properties and roles of the SGCs is ongoing.

Nerve injury is an injury to a nerve. There is no single classification system that can describe all the many variations of nerve injuries. In 1941, Seddon introduced a classification of nerve injuries based on three main types of nerve fiber injury and whether there is continuity of the nerve. Usually, however, nerve injuries are classified in five stages, based on the extent of damage to both the nerve and the surrounding connective tissue, since supporting glial cells may be involved.
Neuroregeneration involves the regrowth or repair of nervous tissues, cells or cell products. Neuroregenerative mechanisms may include generation of new neurons, glia, axons, myelin, or synapses. Neuroregeneration differs between the peripheral nervous system (PNS) and the central nervous system (CNS) by the functional mechanisms involved, especially in the extent and speed of repair. When an axon is damaged, the distal segment undergoes Wallerian degeneration, losing its myelin sheath. The proximal segment can either die by apoptosis or undergo the chromatolytic reaction, which is an attempt at repair. In the CNS, synaptic stripping occurs as glial foot processes invade the dead synapse.
Protective autoimmunity is a condition in which cells of the adaptive immune system contribute to maintenance of the functional integrity of a tissue, or facilitate its repair following an insult. The term ‘protective autoimmunity’ was coined by Prof. Michal Schwartz of the Weizmann Institute of Science (Israel), whose pioneering studies were the first to demonstrate that autoimmune T lymphocytes can have a beneficial role in repair, following an injury to the central nervous system (CNS). Most of the studies on the phenomenon of protective autoimmunity were conducted in experimental settings of various CNS pathologies and thus reside within the scientific discipline of neuroimmunology.

Olfactory ensheathing cells (OECs), also known as olfactory ensheathing glia or olfactory ensheathing glial cells, are a type of macroglia found in the nervous system. They are also known as olfactory Schwann cells, because they ensheath the non-myelinated axons of olfactory neurons in a similar way to which Schwann cells ensheath non-myelinated peripheral neurons. They also share the property of assisting axonal regeneration.
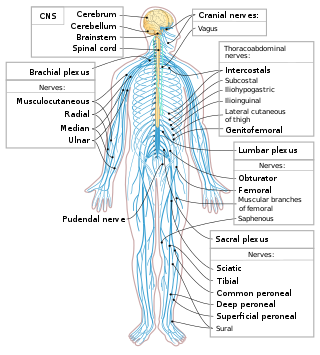
The following diagram is provided as an overview of and topical guide to the human nervous system:
References
- ↑ Juul SE, Anderson DK, Li Y, Christensen RD (January 1998). "Erythropoietin and erythropoietin receptor in the developing human central nervous system". Pediatr. Res. 43 (1): 40–9. doi: 10.1203/00006450-199804001-00243 . PMID 9432111.
- 1 2 Grimm C, Wenzel A, Groszer M, Mayser H, Seeliger M, Samardzija M, Bauer C, Gassmann M, Remé CE (July 2002). "HIF-1-induced erythropoietin in the hypoxic retina protects against light-induced retinal degeneration". Nat. Med. 8 (7): 718–24. doi:10.1038/nm723. PMID 12068288. S2CID 1595426.
- ↑ Morishita E, Masuda S, Nagao M, Yasuda Y, Sasaki R (January 1997). "Erythropoietin receptor is expressed in rat hippocampal and cerebral cortical neurons, and erythropoietin prevents in vitro glutamate-induced neuronal death". Neuroscience. 76 (1): 105–16. doi:10.1016/S0306-4522(96)00306-5. PMID 8971763. S2CID 41374229.
- ↑ Celik M, Gökmen N, Erbayraktar S, Akhisaroglu M, Konakc S, Ulukus C, Genc S, Genc K, Sagiroglu E, Cerami A, Brines M (February 2002). "Erythropoietin prevents motor neuron apoptosis and neurologic disability in experimental spinal cord ischemic injury". Proc. Natl. Acad. Sci. U.S.A. 99 (4): 2258–63. Bibcode:2002PNAS...99.2258C. doi: 10.1073/pnas.042693799 . PMC 122352 . PMID 11854521.
- ↑ Yu X, Shacka JJ, Eells JB, Suarez-Quian C, Przygodzki RM, Beleslin-Cokic B, Lin CS, Nikodem VM, Hempstead B, Flanders KC, Costantini F, Noguchi CT (January 2002). "Erythropoietin receptor signalling is required for normal brain development". Development. 129 (2): 505–16. doi:10.1242/dev.129.2.505. PMID 11807041.
- ↑ Suzuki N, Ohneda O, Takahashi S, Higuchi M, Mukai HY, Nakahata T, Imagawa S, Yamamoto M (October 2002). "Erythroid-specific expression of the erythropoietin receptor rescued its null mutant mice from lethality". Blood. 100 (7): 2279–88. doi:10.1182/blood-2002-01-0124. hdl: 2241/1805 . PMID 12239135. S2CID 35004292.
- ↑ Yuan, R.; Wang, B.; Lu, W.; Maeda, Y.; Dowling, P. (2015). "A Distinct Region in Erythropoietin that Induces Immuno/Inflammatory Modulation and Tissue Protection". Neurotherapeutics. 12 (4): 850–861. doi:10.1007/s13311-015-0379-1. PMC 4604189 . PMID 26271954.
- ↑ "Short Stabilized EPO-Peptide as Side-Effect-Free Therapeutic Agents for Multiple Sclerosis and Acute Brain Trauma". National Center for Advancing Translational Science (NCATS). 2015-03-12.
- ↑ Campana WM, Myers RR (August 2001). "Erythropoietin and erythropoietin receptors in the peripheral nervous system: changes after nerve injury". FASEB J. 15 (10): 1804–6. doi: 10.1096/fj.00-0857fje . PMID 11481236. S2CID 11803744.
- ↑ Bernaudin M, Bellail A, Marti HH, Yvon A, Vivien D, Duchatelle I, Mackenzie ET, Petit E (May 2000). "Neurons and astrocytes express EPO mRNA: oxygen-sensing mechanisms that involve the redox-state of the brain". Glia. 30 (3): 271–8. doi:10.1002/(sici)1098-1136(200005)30:3<271::aid-glia6>3.0.co;2-h. PMID 10756076. S2CID 10131344.
- 1 2 3 4 5 Lykissas MG, Korompilias AV, Vekris MD, Mitsionis GI, Sakellariou E, Beris AE (October 2007). "The role of erythropoietin in central and peripheral nerve injury". Clin Neurol Neurosurg. 109 (8): 639–44. doi:10.1016/j.clineuro.2007.05.013. PMID 17624659. S2CID 22990605.
- ↑ Keswani SC, Buldanlioglu U, Fischer A, Reed N, Polley M, Liang H, Zhou C, Jack C, Leitz GJ, Hoke A (December 2004). "A novel endogenous erythropoietin mediated pathway prevents axonal degeneration". Ann. Neurol. 56 (6): 815–26. doi:10.1002/ana.20285. PMID 15470751. S2CID 22025115.
- ↑ Springborg JB, Ma X, Rochat P, Knudsen GM, Amtorp O, Paulson OB, Juhler M, Olsen NV (February 2002). "A single subcutaneous bolus of erythropoietin normalizes cerebral blood flow autoregulation after subarachnoid haemorrhage in rats". Br. J. Pharmacol. 135 (3): 823–9. doi:10.1038/sj.bjp.0704521. PMC 1573185 . PMID 11834631.
- ↑ Chong ZZ, Kang JQ, Maiese K (May 2002). "Hematopoietic factor erythropoietin fosters neuroprotection through novel signal transduction cascades". J. Cereb. Blood Flow Metab. 22 (5): 503–14. doi: 10.1097/00004647-200205000-00001 . PMID 11973422.
- ↑ Sirén AL, Fratelli M, Brines M, Goemans C, Casagrande S, Lewczuk P, Keenan S, Gleiter C, Pasquali C, Capobianco A, Mennini T, Heumann R, Cerami A, Ehrenreich H, Ghezzi P (March 2001). "Erythropoietin prevents neuronal apoptosis after cerebral ischemia and metabolic stress". Proc. Natl. Acad. Sci. U.S.A. 98 (7): 4044–9. Bibcode:2001PNAS...98.4044S. doi: 10.1073/pnas.051606598 . PMC 31176 . PMID 11259643.
- ↑ Digicaylioglu M, Lipton SA (August 2001). "Erythropoietin-mediated neuroprotection involves cross-talk between Jak2 and NF-kappaB signalling cascades". Nature. 412 (6847): 641–7. Bibcode:2001Natur.412..641D. doi:10.1038/35088074. PMID 11493922. S2CID 4346187.
- ↑ Yamamoto Y, Gaynor RB (July 2001). "Role of the NF-kappaB pathway in the pathogenesis of human disease states". Curr. Mol. Med. 1 (3): 287–96. doi:10.2174/1566524013363816. PMID 11899077.
- ↑ Campana WM, Myers RR (September 2003). "Exogenous erythropoietin protects against dorsal root ganglion apoptosis and pain following peripheral nerve injury". Eur. J. Neurosci. 18 (6): 1497–506. doi:10.1046/j.1460-9568.2003.02875.x. PMID 14511329. S2CID 8533004.
- 1 2 McPherson RJ, Juul SE (February 2008). "Recent trends in erythropoietin-mediated neuroprotection". Int. J. Dev. Neurosci. 26 (1): 103–11. doi:10.1016/j.ijdevneu.2007.08.012. PMC 2312376 . PMID 17936539.
- ↑ Gonzalez FF, McQuillen P, Mu D, Chang Y, Wendland M, Vexler Z, Ferriero DM (2007). "Erythropoietin enhances long-term neuroprotection and neurogenesis in neonatal stroke". Dev. Neurosci. 29 (4–5): 321–30. doi: 10.1159/000105473 . PMID 17762200.
- ↑ Mogensen J, Jensen C, Kingod SC, Hansen A, Larsen JA, Malá H (January 2008). "Erythropoietin improves spatial delayed alternation in a T-maze in fimbria-fornix transected rats". Behav. Brain Res. 186 (2): 215–21. doi:10.1016/j.bbr.2007.08.009. PMID 17888525. S2CID 22974206.
- ↑ Xue YQ, Zhao LR, Guo WP, Duan WM (May 2007). "Intrastriatal administration of erythropoietin protects dopaminergic neurons and improves neurobehavioral outcome in a rat model of Parkinson's disease". Neuroscience. 146 (3): 1245–58. doi:10.1016/j.neuroscience.2007.02.004. PMID 17363174. S2CID 10775364.
- 1 2 Brines M (September 2002). "What evidence supports use of erythropoietin as a novel neurotherapeutic?". Oncology (Williston Park, N.Y.). 16 (9 Suppl 10): 79–89. PMID 12380958.
- ↑ Tsai PT, Ohab JJ, Kertesz N, Groszer M, Matter C, Gao J, Liu X, Wu H, Carmichael ST (2006). "A critical role of erythropoietin receptor in neurogenesis and post-stroke recovery". J. Neurosci. 26 (4): 1269–74. doi:10.1523/JNEUROSCI.4480-05.2006. PMC 6674578 . PMID 16436614.
- ↑ King CE, Rodger J, Bartlett C, Esmaili T, Dunlop SA, Beazley LD (May 2007). "Erythropoietin is both neuroprotective and neuroregenerative following optic nerve transection". Exp. Neurol. 205 (1): 48–55. doi:10.1016/j.expneurol.2007.01.017. PMID 17328893. S2CID 43344936.
- ↑ Zhong L, Bradley J, Schubert W, Ahmed E, Adamis AP, Shima DT, Robinson GS, Ng YS (March 2007). "Erythropoietin promotes survival of retinal ganglion cells in DBA/2J glaucoma mice". Invest. Ophthalmol. Vis. Sci. 48 (3): 1212–8. doi:10.1167/iovs.06-0757. PMID 17325165.
Erythropioetin is a very potent molecule (One Unit ~ 10 ng)and is known to be extremely toxic in higher doses. Since, such a tight control of expression is impossible, genetherapy with viral vectors is not an option