Related Research Articles

Ungulates are members of the diverse clade Euungulata which primarily consists of large mammals with hooves. Once part of the clade "Ungulata" along with the clade Paenungulata, "Ungulata" has since been determined to be a polyphyletic and thereby invalid clade based on molecular data. As a result, true ungulates had since been reclassified to the newer clade Euungulata in 2001 within the clade Laurasiatheria while Paenungulata has been reclassified to a distant clade Afrotheria. Living ungulates are divided into two orders: the odd-toed ungulates (Perissodactyla) including horses, rhinoceroses, and tapirs; and even-toed ungulates (Artiodactyla) such as cattle, pigs, giraffes, camels, sheep, deer, and hippopotamuses. Cetaceans such as whales, dolphins, and porpoises are also classified as even-toed ungulates, although they do not have hooves. Most terrestrial ungulates use the hoofed tips of their toes to support their body weight while standing or moving. Two other orders of ungulates, Notoungulata and Litopterna, both native to South America, became extinct at the end of the Pleistocene, around 12,000 years ago.

The molars or molar teeth are large, flat teeth at the back of the mouth. They are more developed in mammals. They are used primarily to grind food during chewing. The name molar derives from Latin, molaris dens, meaning "millstone tooth", from mola, millstone and dens, tooth. Molars show a great deal of diversity in size and shape across mammal groups. The third molar of humans is sometimes vestigial.
Lavanify is a mammalian genus from the late Cretaceous of Madagascar. The only species, L. miolaka, is known from two isolated teeth, one of which is damaged. The teeth were collected in 1995–1996 and described in 1997. The animal is classified as a member of Gondwanatheria, an enigmatic extinct group with unclear phylogenetic relationships, and within Gondwanatheria as a member of the family Sudamericidae. Lavanify is most closely related to the Indian Bharattherium; the South American Sudamerica and Gondwanatherium are more distantly related. Gondwanatheres probably ate hard plant material.

Elasmotherium is an extinct genus of large rhinoceros endemic to Eurasia during Late Miocene through to the Late Pleistocene, with the youngest reliable dates around 39,000 years ago. It was the last surviving member of Elasmotheriinae, a distinctive group of rhinoceroses separate from the group that contains living rhinoceros (Rhinocerotinae). The two groups are estimated to have split at least 35 million years ago according to fossils and molecular evidence.
Hypsodont is a pattern of dentition with high-crowned teeth and enamel extending past the gum line, providing extra material for wear and tear. Some examples of animals with hypsodont dentition are cows and horses; all animals that feed on gritty, fibrous material. The opposite condition is called brachydont.
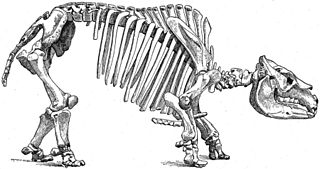
Notoungulata is an extinct order of mammalian ungulates that inhabited South America from the early Paleocene to the Holocene, living from approximately 61 million to 11,000 years ago. Notoungulates were morphologically diverse, with forms resembling animals as disparate as rabbits and rhinoceroses. Notoungulata are the largest group of South American native ungulates, with over 150 genera in 14 families having been described, divided into two major subgroupings, Typotheria and Toxodontia. Notoungulates first diversified during the Eocene. Their diversity declined during the Late Neogene, with only the large toxodontids persisting until the end of the Pleistocene, perishing as part of the Quaternary extinction event among with most other large mammals in the Americas. Collagen analysis suggests that notoungulates are closely related to litopterns, another group of South American ungulates, and their closest living relatives being perissodactyls, including rhinoceroses, tapirs and equines. However their relationships to other South American ungulates are uncertain. Several groups of notoungulates separately evolved ever-growing cheek teeth.

Toxodontia is a suborder of the meridiungulate order Notoungulata. Most of the members of the five included families, including the largest notoungulates, share several dental, auditory and tarsal specializations. The group is named after Toxodon, the first example of the group to be discovered by science.
The fossil Tinguiririca fauna, entombed in volcanic mudflows and ash layers at the onset of the Oligocene, about 33-31.5 million years ago, represents a unique snapshot of the history of South America's endemic fauna, which was extinguished when the former island continent was joined to North America by the rising Isthmus of Panama. The fossil-bearing sedimentary layers of the Abanico Formation were first discovered in the valley of the Tinguiririca River, high in the Andes of central Chile. The faunal assemblage lends its name to the Tinguirirican stage in the South American land mammal age (SALMA) classification.

Hadrosaurids, also commonly referred to as duck-billed dinosaurs or hadrosaurs, were large terrestrial herbivores. The diet of hadrosaurid dinosaurs remains a subject of debate among paleontologists, especially regarding whether hadrosaurids were grazers who fed on vegetation close to the ground, or browsers who ate higher-growing leaves and twigs. Preserved stomach content findings have indicated they may have been browsers, whereas other studies into jaw movements indicate they may have been grazers.
Mesowear is a method, used in different branches and fields of biology. This method can apply to both extant and extinct animals, according to the scope of the study. Mesowear is based on studying an animal's tooth wearing fingerprint. In brief, each animal has special feeding habits, which cause unique tooth wearing. Rough feeds cause serious tooth abrasion, while smooth one triggers moderate abrasion, so browsers have teeth with moderate abrasion and grazers have teeth with rough abrasion. Scoring systems can quantify tooth abrasion observations and ease comparisons between individuals.
Bharattherium is a mammal that lived in India during the Maastrichtian and possibly the Paleocene. The genus has a single species, Bharattherium bonapartei. It is part of the gondwanathere family Sudamericidae, which is also found in Madagascar and South America during the latest Cretaceous. The first fossil of Bharattherium was discovered in 1989 and published in 1997, but the animal was not named until 2007, when two teams independently named the animal Bharattherium bonapartei and Dakshina jederi. The latter name is now a synonym. Bharattherium is known from a total of eight isolated fossil teeth, including one incisor and seven molariforms.

Piauhytherium is an extinct genus of herbivorous notoungulate mammal of the family Toxodontidae. It lived during the Late Pleistocene; fossils have been found in Brazil. The only known species is Piauhytherium capivarae.

Christine Marie Janis is a British palaeontologist who specialises in mammals. She is currently based at the University of Bristol.

Prosotherium is an extinct genus of hegetotheriid notoungulate. It lived during the Late Oligocene, and its fossilized remains were found in South America.
Eohyrax is an extinct genus of Notoungulate, belonging to the suborder Typotheria. It lived during the Middle Eocene, and its remains were discovered in South America.
Pseudhyrax is an extinct genus of archaeohyracid notoungulate. It lived from the Late Eocene to the Early Oligocene, of what is now South America.

Eomorphippus is an extinct genus of notohippid notoungulate that lived from the Late Eocene to the Early Oligocene in what is today South America.

Morphippus is an extinct genus of notohippid notoungulate that lived during the Middle to Late Oligocene in what is now South America.
Archaeopithecus is an extinct genus of Notoungulate, belonging to the suborder Typotheria. It lived during the Middle Eocene, in what is today Argentina.
Rosendo is an extinct genus of notohippid notoungulates that lived during the Early Oligocene in what is now Argentina and Chile. Fossils of this genus have been found in the Sarmiento Formation and the Abanico Formations of Argentina and Chile.
References
- 1 2 3 Jardine, Phillip E.; Janis, Christine M.; Sahney, Sarda; Benton, Michael J. (2012). "Grit not grass: Concordant patterns of early origin of hypsodonty in Great Plains ungulates and Glires". Palaeogeography, Palaeoclimatology, Palaeoecology. 365–366: 1–10. Bibcode:2012PPP...365....1J. doi:10.1016/j.palaeo.2012.09.001.
- ↑ Stirton, R. A. (1947). "Observations on Evolutionary Rates in Hypsodonty". Evolution. 1 (1–2): 32–41. doi: 10.1111/j.1558-5646.1947.tb02711.x . S2CID 87532435.
- ↑ Caroline A. E. Strömberg (2006). "Evolution of hypsodonty in equids: testing a hypothesis of adaptation" (PDF). Paleobiology. 32 (2): 236–258. doi:10.1666/0094-8373(2006)32[236:eohiet]2.0.co;2. S2CID 12338144.
- 1 2 Mendoza, M.; Palmqvist, P. (February 2008). "Hypsodonty in ungulates: an adaptation for grass consumption or for foraging in open habitat?" (PDF). Journal of Zoology. 274 (2): 134–142. doi:10.1111/j.1469-7998.2007.00365.x.
- 1 2 3 Billet, Blondel, and Muizon (2009), "Dental microwear analysis of notoungulates (Mammalia) from Salla (Late Oligocene, Bolivia) and discussion on their precocious hypsodonty", Palaeogeography, Palaeoclimatology, Palaeoecology, 274 (1–2): 114–124, Bibcode:2009PPP...274..114B, doi:10.1016/j.palaeo.2009.01.004
{{citation}}: CS1 maint: multiple names: authors list (link) - ↑ "Hadrosaur chowdown — grind, grind, grind", Associated Press, 2009
- ↑ Flynn, John J.; Wyss, André R.; Charrier, Reynaldo (May 2007). "South America's Missing Mammals". Scientific American. 296 (5): 68–75. Bibcode:2007SciAm.296e..68F. doi:10.1038/scientificamerican0507-68. PMID 17500416.
- ↑ Kwan, Paul W.L. (2007). "Digestive system I" (PDF). Tufts University.