Related Research Articles

A herbivore is an animal anatomically and physiologically adapted to eating plant material, for example foliage or marine algae, for the main component of its diet. As a result of their plant diet, herbivorous animals typically have mouthparts adapted to rasping or grinding. Horses and other herbivores have wide flat teeth that are adapted to grinding grass, tree bark, and other tough plant material.

A lima bean, also commonly known as the butter bean, sieva bean, double bean or Madagascar bean is a legume grown for its edible seeds or beans.
Chemical ecology is the study of chemically mediated interactions between living organisms, and the effects of those interactions on the demography, behavior and ultimately evolution of the organisms involved. It is thus a vast and highly interdisciplinary field. Chemical ecologists seek to identify the specific molecules that function as signals mediating community or ecosystem processes and to understand the evolution of these signals. The substances that serve in such roles are typically small, readily-diffusible organic molecules, but can also include larger molecules and small peptides.

Jasmonate (JA) and its derivatives are lipid-based plant hormones that regulate a wide range of processes in plants, ranging from growth and photosynthesis to reproductive development. In particular, JAs are critical for plant defense against herbivory and plant responses to poor environmental conditions and other kinds of abiotic and biotic challenges. Some JAs can also be released as volatile organic compounds (VOCs) to permit communication between plants in anticipation of mutual dangers.

Phytoalexins are antimicrobial substances, some of which are antioxidative as well. They are defined, not by their having any particular chemical structure or character, but by the fact that they are defensively synthesized de novo by plants that produce the compounds rapidly at sites of pathogen infection. In general phytoalexins are broad spectrum inhibitors; they are chemically diverse, and different chemical classes of compounds are characteristic of particular plant taxa. Phytoalexins tend to fall into several chemical classes, including terpenoids, glycosteroids, and alkaloids; however the term applies to any phytochemicals that are induced by microbial infection.

Methyl jasmonate is a volatile organic compound used in plant defense and many diverse developmental pathways such as seed germination, root growth, flowering, fruit ripening, and senescence. Methyl jasmonate is derived from jasmonic acid and the reaction is catalyzed by S-adenosyl-L-methionine:jasmonic acid carboxyl methyltransferase.
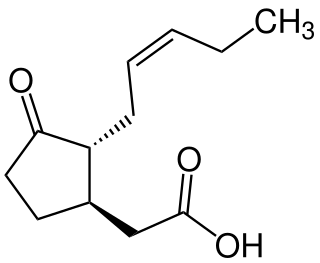
Jasmonic acid (JA) is an organic compound found in several plants including jasmine. The molecule is a member of the jasmonate class of plant hormones. It is biosynthesized from linolenic acid by the octadecanoid pathway. It was first isolated in 1957 as the methyl ester of jasmonic acid by the Swiss chemist Édouard Demole and his colleagues.

Plant defense against herbivory or host-plant resistance (HPR) is a range of adaptations evolved by plants which improve their survival and reproduction by reducing the impact of herbivores. Plants can sense being touched, and they can use several strategies to defend against damage caused by herbivores. Many plants produce secondary metabolites, known as allelochemicals, that influence the behavior, growth, or survival of herbivores. These chemical defenses can act as repellents or toxins to herbivores or reduce plant digestibility. Another defensive strategy of plants is changing their attractiveness. To prevent overconsumption by large herbivores, plants alter their appearance by changing their size or quality, reducing the rate at which they are consumed.
Herbivores are dependent on plants for food, and have coevolved mechanisms to obtain this food despite the evolution of a diverse arsenal of plant defenses against herbivory. Herbivore adaptations to plant defense have been likened to "offensive traits" and consist of those traits that allow for increased feeding and use of a host. Plants, on the other hand, protect their resources for use in growth and reproduction, by limiting the ability of herbivores to eat them. Relationships between herbivores and their host plants often results in reciprocal evolutionary change. When a herbivore eats a plant it selects for plants that can mount a defensive response, whether the response is incorporated biochemically or physically, or induced as a counterattack. In cases where this relationship demonstrates "specificity", and "reciprocity", the species are thought to have coevolved. The escape and radiation mechanisms for coevolution, presents the idea that adaptations in herbivores and their host plants, has been the driving force behind speciation. The coevolution that occurs between plants and herbivores that ultimately results in the speciation of both can be further explained by the Red Queen hypothesis. This hypothesis states that competitive success and failure evolve back and forth through organizational learning. The act of an organism facing competition with another organism ultimately leads to an increase in the organism's performance due to selection. This increase in competitive success then forces the competing organism to increase its performance through selection as well, thus creating an "arms race" between the two species. Herbivores evolve due to plant defenses because plants must increase their competitive performance first due to herbivore competitive success.
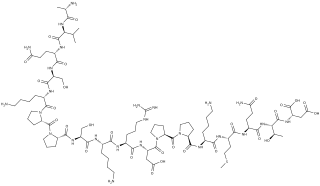
Systemin is a plant peptide hormone involved in the wound response in the family Solanaceae. It was the first plant hormone that was proven to be a peptide having been isolated from tomato leaves in 1991 by a group led by Clarence A. Ryan. Since then, other peptides with similar functions have been identified in tomato and outside of the Solanaceae. Hydroxyproline-rich glycopeptides were found in tobacco in 2001 and AtPeps were found in Arabidopsis thaliana in 2006. Their precursors are found both in the cytoplasm and cell walls of plant cells, upon insect damage, the precursors are processed to produce one or more mature peptides. The receptor for systemin was first thought to be the same as the brassinolide receptor but this is now uncertain. The signal transduction processes that occur after the peptides bind are similar to the cytokine-mediated inflammatory immune response in animals. Early experiments showed that systemin travelled around the plant after insects had damaged the plant, activating systemic acquired resistance, now it is thought that it increases the production of jasmonic acid causing the same result. The main function of systemins is to coordinate defensive responses against insect herbivores but they also affect plant development. Systemin induces the production of protease inhibitors which protect against insect herbivores, other peptides activate defensins and modify root growth. They have also been shown to affect plants' responses to salt stress and UV radiation. AtPEPs have been shown to affect resistance against oomycetes and may allow A. thaliana to distinguish between different pathogens. In Nicotiana attenuata, some of the peptides have stopped being involved in defensive roles and instead affect flower morphology.
Biotic stress is stress that occurs as a result of damage done to an organism by other living organisms, such as bacteria, viruses, fungi, parasites, beneficial and harmful insects, weeds, and cultivated or native plants. It is different from abiotic stress, which is the negative impact of non-living factors on the organisms such as temperature, sunlight, wind, salinity, flooding and drought. The types of biotic stresses imposed on an organism depend the climate where it lives as well as the species' ability to resist particular stresses. Biotic stress remains a broadly defined term and those who study it face many challenges, such as the greater difficulty in controlling biotic stresses in an experimental context compared to abiotic stress.

Nicotiana attenuata is a species of wild tobacco known by the common name coyote tobacco. It is native to western North America from British Columbia to Texas and northern Mexico, where it grows in many types of habitat. It is a glandular and sparsely hairy annual herb exceeding a meter in maximum height. The leaf blades may be 10 centimetres (4 in) long, the lower ones oval and the upper narrower in shape, and are borne on petioles. The inflorescence bears several flowers with pinkish or greenish white tubular throats 2 to 3 centimetres long, their bases enclosed in pointed sepals. The flower face has five mostly white lobes. The fruit is a capsule about 1 centimetre long.
Tolerance is the ability of plants to mitigate the negative fitness effects caused by herbivory. It is one of the general plant defense strategies against herbivores, the other being resistance, which is the ability of plants to prevent damage. Plant defense strategies play important roles in the survival of plants as they are fed upon by many different types of herbivores, especially insects, which may impose negative fitness effects. Damage can occur in almost any part of the plants, including the roots, stems, leaves, flowers and seeds. In response to herbivory, plants have evolved a wide variety of defense mechanisms and although relatively less studied than resistance strategies, tolerance traits play a major role in plant defense.
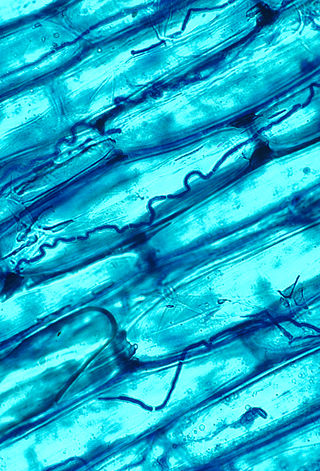
Plant use of endophytic fungi in defense occurs when endophytic fungi, which live symbiotically with the majority of plants by entering their cells, are utilized as an indirect defense against herbivores. In exchange for carbohydrate energy resources, the fungus provides benefits to the plant which can include increased water or nutrient uptake and protection from phytophagous insects, birds or mammals. Once associated, the fungi alter nutrient content of the plant and enhance or begin production of secondary metabolites. The change in chemical composition acts to deter herbivory by insects, grazing by ungulates and/or oviposition by adult insects. Endophyte-mediated defense can also be effective against pathogens and non-herbivory damage.
Wilhelm Boland is a German chemist.

Ian Thomas Baldwin is an American ecologist.

Escape and radiate coevolution is a hypothesis proposing that a coevolutionary 'arms-race' between primary producers and their consumers contributes to the diversification of species by accelerating speciation rates. The hypothesized process involves the evolution of novel defenses in the host, allowing it to "escape" and then "radiate" into differing species.

Tritrophic interactions in plant defense against herbivory describe the ecological impacts of three trophic levels on each other: the plant, the herbivore, and its natural enemies. They may also be called multitrophic interactions when further trophic levels, such as soil microbes, endophytes, or hyperparasitoids are considered. Tritrophic interactions join pollination and seed dispersal as vital biological functions which plants perform via cooperation with animals.
Plants are constantly exposed to different stresses that result in wounding. Plants have adapted to defend themselves against wounding events, like herbivore attacks or environmental stresses. There are many defense mechanisms that plants rely on to help fight off pathogens and subsequent infections. Wounding responses can be local, like the deposition of callose, and others are systemic, which involve a variety of hormones like jasmonic acid and abscisic acid.
Cryptic mimicry is observed in animals as well as plants. In animals, this may involve nocturnality, camouflage, subterranean lifestyle, and mimicry. Generally, plant herbivores are visually oriented. So a mimicking plant should strongly resemble its host; this can be done through visual and/or textural change. Previous criteria for mimicry include similarity of leaf dimensions, leaf presentation, and intermodal distances between the host and mimicking plant.
References
- 1 2 Karban R, Baldwin IT. Induced responses to herbivory. Chicago: Chicago University Press; 1997.
- ↑ Chen MS. Inducible direct plant defence against insect herbivores: a review. Insect Science. 2008;15:101-14.
- ↑ Dicke M, van Poecke RMP, de Boer JG. Inducible indirect defence of plants: from mechanisms to ecological functions. Basic and Applied Ecology. 2003;4:27-42.
- ↑ Gatehouse JA. Plant resistance towards insect herbivores: a dynamic interaction. New Phytologist. 2002;156:145-69.
- 1 2 Karban R, Agrawal AA, Mangel M. The benefits of induced defenses against herbivores. Ecology. 1997;78:1351-5.
- 1 2 Shelton AL. Variation in chemical defenses of plants may improve the effectiveness of defense. Evolutionary Ecological Research 2004;6:709-26.
- ↑ Agrawal AA. Induced responses to herbivory in wild radish: effects on several herbivores and plant fitness. Ecology. 1999;80:1713-23.
- ↑ Steppuhn A, Gase K, Krock B, Halitschke R, Baldwin IT. Nicotine's Defensive Function in Nature. PLoS Biology. 2004 August 01, 2004;2(8):e217.
- ↑ Preisser EL, Gibson SE, Adler LS, Lewis EE. Underground herbivory and the costs of constitutive defense in tobacco. Acta Oecol-Int J Ecol. 2007;31:210-5.
- ↑ Milewski, A.V., T.P. Young and D. Madden. 1991. Thorns as induced defenses: experimental evidence. Oecologia 86:70-75.
- ↑ Young, T.P., M.L. Stanton and C. Christian. 2003. Effects of natural and simulated herbivory on spine lengths of Acacia drepanolobium in Kenya. Oikos 101:171-179.
- ↑ Huntzinger, P.M., R. Karban, T.P. Young and T.M. Palmer. 2004. Relaxation of induced indirect defenses of acacias following exclusion of mammalian herbivores. Ecology 85:609-614.
- ↑ Mole S. Trade-offs and constraints in plant-herbivore defense theory: a life history perspective. Okios. 1994;71:3-12.
- ↑ Strauss SY, Rudgers JA, Lau JA, Irwin RE. Direct and ecological costs of resistance to herbivory. Trends in Ecology & Evolution. 2002;17:278-85.
- 1 2 Heil M, Baldwin IT. Fitness costs of induced resistance: emerging experimental support for a slippery concept. Trends in Plant Science. 2002;7:61-7.
- ↑ Baldwin IT, Ohnmeiss TE. Swords into plowghshares? Nicotiana sylvestris does not use nicotine as a nitrogen source under nitrogen-limited growth Oecologia. 1994;98:385-92.
- ↑ Herms DA, Mattson WJ. The dilemma of plants: to grow or to defend. Quarterly Review of Biology. 1992;67:283-335.
- ↑ Moor JP, Taylor JE, Paul ND, Whittaker JB. Reduced leaf expansion as a cost of systematic induced resistance to herbivory. Functional Ecology. 2003;17:75-81.
- ↑ Ataide, Livia M. S.; Pappas, Maria L.; Schimmel, Bernardus C. J.; Lopez-Orenes, Antonio; Alba, Juan M.; Duarte, Marcus V. A.; Pallini, Angelo; Schuurink, Robert C.; Kant, Merijn R. (2016-11-01). "Induced plant-defenses suppress herbivore reproduction but also constrain predation of their offspring". Plant Science. 252: 300–310. doi: 10.1016/j.plantsci.2016.08.004 . ISSN 0168-9452.
- ↑ Redman AM, Cipollini DF, Schultz JC. Fitness cost of jasmonic acid-induced defense in tomato, Lycopersicon esculentum. Oecologia. 2001;126:380-5.
- ↑ Giovannoni JJ. Genetic regulation of fruit development and ripening. Plant Cell. 2004;16:S170-S80.
- ↑ Boyd, Jade (2012). "A bit touchy: Plants' insect defenses activated by touch". Rice University. http://news.rice.edu/2012/04/09/a-bit-touchy-plants-insect-defenses-activated-by-touch/
- 1 2 Korth KL, Dixon RA. Evidence for chewing insect-specific molecular events distinct from a general wound response in leaves. Plant Physiology. 1997;115:1299-305.
- ↑ Tamiru (2017). Bruce TJA, Richter A, Woodcock CM, Midega CAO, Degenhardt J, Kelemu S, Pickett JA, Khan ZRA (2017) A maize landrace that emits defense volatiles in response toherbivore eggs possesses a strongly inducible terpene synthase gene. Ecology and Evolution 7: 2835-2845
- ↑ Walling LL. The myriad plant responses to herbivores. Journal of Plant Growth Regulation. 2000;19:195-216.
- ↑ Hermsmeier D, Schittko U, Baldwin IT. Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. 1. Large-scale changes in the accumulation of growth- and defense-related plant mRNAs. Plant Physiology. 2001;125:1875-87.
- ↑ Schenk JM, Kazan K, Wilson I, Anderson JP, Richmond T, Somerville SC, et al. Coordinated plant defence responses in Arabidopsis revealed by microarray analysis. Proceedings of the National Academy of Sciences, USA. 2000;97:11655-60.
- ↑ Ryan CA. The systemin signaling pathway: differential activation of plant defensive genes. Biochimica et Biophysica Acta-Protein Structure and Molecular Enzymology. 2000;1477(1-2):112-21.
- ↑ Frost CJ, Appel HM, Carlson JE, Moraes CMD, Mescher MC, Schultz JC. Within-plant signalling via volatiles overcomes vascular constraints on systemic signalling and primes responses against herbivores. Ecology Letters. 2007;10(6):490-8.