
A herbivore is an animal anatomically and physiologically adapted to eating plant material, for example foliage or marine algae, for the main component of its diet. As a result of their plant diet, herbivorous animals typically have mouthparts adapted to rasping or grinding. Horses and other herbivores have wide flat teeth that are adapted to grinding grass, tree bark, and other tough plant material.

Datura stramonium, known by the common names thorn apple, jimsonweed, devil's snare, or devil's trumpet, is a poisonous flowering plant of the nightshade family Solanaceae. It is a species belonging to the Datura genus and Daturae tribe. Its likely origin was in Central America, and it has been introduced in many world regions. It is an aggressive invasive weed in temperate climates and tropical climates across the world. D. stramonium has frequently been employed in traditional medicine to treat a variety of ailments. It has also been used as a hallucinogen, taken entheogenically to cause intense, sacred or occult visions. It is unlikely ever to become a major drug of abuse owing to effects upon both mind and body frequently perceived as being highly unpleasant, giving rise to a state of profound and long-lasting disorientation or delirium with a potentially fatal outcome. It contains tropane alkaloids which are responsible for the psychoactive effects, and may be severely toxic.

Plant defense against herbivory or host-plant resistance (HPR) is a range of adaptations evolved by plants which improve their survival and reproduction by reducing the impact of herbivores. Plants can sense being touched, and they can use several strategies to defend against damage caused by herbivores. Many plants produce secondary metabolites, known as allelochemicals, that influence the behavior, growth, or survival of herbivores. These chemical defenses can act as repellents or toxins to herbivores or reduce plant digestibility. Another defensive strategy of plants is changing their attractiveness. To prevent overconsumption by large herbivores, plants alter their appearance by changing their size or quality, reducing the rate at which they are consumed.
Ecological facilitation or probiosis describes species interactions that benefit at least one of the participants and cause harm to neither. Facilitations can be categorized as mutualisms, in which both species benefit, or commensalisms, in which one species benefits and the other is unaffected. This article addresses both the mechanisms of facilitation and the increasing information available concerning the impacts of facilitation on community ecology.
Herbivores are dependent on plants for food, and have coevolved mechanisms to obtain this food despite the evolution of a diverse arsenal of plant defenses against herbivory. Herbivore adaptations to plant defense have been likened to "offensive traits" and consist of those traits that allow for increased feeding and use of a host. Plants, on the other hand, protect their resources for use in growth and reproduction, by limiting the ability of herbivores to eat them. Relationships between herbivores and their host plants often results in reciprocal evolutionary change. When a herbivore eats a plant it selects for plants that can mount a defensive response, whether the response is incorporated biochemically or physically, or induced as a counterattack. In cases where this relationship demonstrates "specificity", and "reciprocity", the species are thought to have coevolved. The escape and radiation mechanisms for coevolution, presents the idea that adaptations in herbivores and their host plants, has been the driving force behind speciation. The coevolution that occurs between plants and herbivores that ultimately results in the speciation of both can be further explained by the Red Queen hypothesis. This hypothesis states that competitive success and failure evolve back and forth through organizational learning. The act of an organism facing competition with another organism ultimately leads to an increase in the organism's performance due to selection. This increase in competitive success then forces the competing organism to increase its performance through selection as well, thus creating an "arms race" between the two species. Herbivores evolve due to plant defenses because plants must increase their competitive performance first due to herbivore competitive success.
The evolution of increased competitive ability (EICA) hypothesis was first proposed by Bernd Blossey and Rolf Nötzold in 1995 as a way to explain the success of invasive, non-indigenous species. Observing that:
- there is usually a lag period between the time of introduction of an invasive species and the point at which it is considered invasive, and
- invasive plants seem to be more virulent in habitats into which they have been introduced, the scientists presumed a sort of naturalization through modification for non-indigenous plants.
Plants and herbivores have co-evolved together for 350 million years. Plants have evolved many defense mechanisms against insect herbivory. Such defenses can be broadly classified into two categories: (1) permanent, constitutive defenses, and (2) temporary, inducible defenses. Both types are achieved through similar means but differ in that constitutive defenses are present before an herbivore attacks, while induced defenses are activated only when attacks occur. In addition to constitutive defenses, initiation of specific defense responses to herbivory is an important strategy for plant persistence and survival.
Evan Siemann is a professor in the Biosciences Department at Rice University in Houston, Texas. He received his AB from Cornell University in 1990 and his PhD from the University of Minnesota in 1997. The focus of his research has been investigating how local environmental factors interact with post-invasion adaptation to determine the likelihood and severity of Chinese tallow tree invasions into East Texas coastal prairie, mesic forests, and floodplain forests. The results of this research have been highlighted in Science Daily, Environmental News Service, and The Sciences. He has also recently begun to explore the ecosystem level impacts of exotic tree invasions into coastal prairies. His research group is also engaged in a number of applied research projects related to controlling exotic plant and animal invasions into Texas ecosystems.
Biotic stress is stress that occurs as a result of damage done to an organism by other living organisms, such as bacteria, viruses, fungi, parasites, beneficial and harmful insects, weeds, and cultivated or native plants. It is different from abiotic stress, which is the negative impact of non-living factors on the organisms such as temperature, sunlight, wind, salinity, flooding and drought. The types of biotic stresses imposed on an organism depend the climate where it lives as well as the species' ability to resist particular stresses. Biotic stress remains a broadly defined term and those who study it face many challenges, such as the greater difficulty in controlling biotic stresses in an experimental context compared to abiotic stress.

Plant ecology is a subdiscipline of ecology that studies the distribution and abundance of plants, the effects of environmental factors upon the abundance of plants, and the interactions among plants and between plants and other organisms. Examples of these are the distribution of temperate deciduous forests in North America, the effects of drought or flooding upon plant survival, and competition among desert plants for water, or effects of herds of grazing animals upon the composition of grasslands.

In plant morphology, thorns, spines, and prickles, and in general spinose structures, are hard, rigid extensions or modifications of leaves, roots, stems, or buds with sharp, stiff ends, and generally serve the same function: physically defending plants against herbivory.
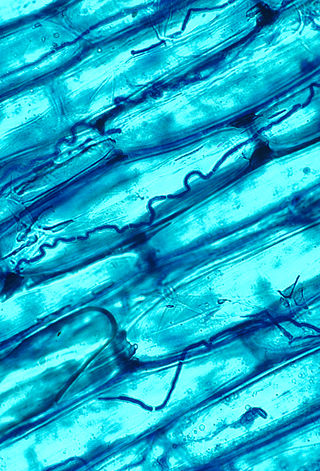
Plant use of endophytic fungi in defense occurs when endophytic fungi, which live symbiotically with the majority of plants by entering their cells, are utilized as an indirect defense against herbivores. In exchange for carbohydrate energy resources, the fungus provides benefits to the plant which can include increased water or nutrient uptake and protection from phytophagous insects, birds or mammals. Once associated, the fungi alter nutrient content of the plant and enhance or begin production of secondary metabolites. The change in chemical composition acts to deter herbivory by insects, grazing by ungulates and/or oviposition by adult insects. Endophyte-mediated defense can also be effective against pathogens and non-herbivory damage.

Community genetics is a recently emerged field in biology that fuses elements of community ecology, evolutionary biology, and molecular and quantitative genetics. Antonovics first articulated the vision for such a field, and Whitham et al. formalized its definition as "The study of the genetic interactions that occur between species and their abiotic environment in complex communities." The field aims to bridge the gaps in the study of evolution and ecology, within the multivariate community context in which ecological and evolutionary features are embedded. The documentary movie A Thousand Invisible Cords provides an introduction to the field and its implications.

Escape and radiate coevolution is a hypothesis proposing that a coevolutionary 'arms-race' between primary producers and their consumers contributes to the diversification of species by accelerating speciation rates. The hypothesized process involves the evolution of novel defenses in the host, allowing it to "escape" and then "radiate" into differing species.

Tritrophic interactions in plant defense against herbivory describe the ecological impacts of three trophic levels on each other: the plant, the herbivore, and its natural enemies. They may also be called multitrophic interactions when further trophic levels, such as soil microbes, endophytes, or hyperparasitoids are considered. Tritrophic interactions join pollination and seed dispersal as vital biological functions which plants perform via cooperation with animals.
Phyllis Dewing Coley is a Biology professor currently teaching at the University of Utah. In 1996 she received the University's Distinguished Research Award. She has been a research associate at the Smithsonian Tropical Research Institute since 1995. In 2023, she was elected to the National Academy of Sciences.

Tecia solanivora, the Central American potato tuberworm, Guatemalan tuber moth, or Guatemalan potato moth, is a moth in the family Gelechiidae. It was described by Povolný in 1973. It is found from Guatemala, through Central America to South America, where it has been recorded from Colombia, Ecuador, Venezuela and Peru. It has also been introduced on the Canary Islands and mainland Spain.
The enemy release hypothesis is among the most widely proposed explanations for the dominance of exotic invasive species. In its native range, a species has co-evolved with pathogens, parasites and predators that limit its population. When it arrives in a new territory, it leaves these old enemies behind, while those in its introduced range are less effective at constraining them. The result is sometimes rampant growth that threatens native species and ecosystems.
Herbivores' effects on plant diversity vary across environmental changes. Herbivores could increase plant diversity or decrease plant diversity. Loss of plant diversity due to climate change can also affect herbivore and plant community relationships
Cryptic mimicry is observed in animals as well as plants. In animals, this may involve nocturnality, camouflage, subterranean lifestyle, and mimicry. Generally, plant herbivores are visually oriented. So a mimicking plant should strongly resemble its host; this can be done through visual and/or textural change. Previous criteria for mimicry include similarity of leaf dimensions, leaf presentation, and intermodal distances between the host and mimicking plant.
