Etymology

The toxin was named ophanin after the snake whose venom it is derived from, the King Cobra ( Ophiophagus hannah ). [1]
Ophanin is a toxin found in the venom of the King Cobra ( Ophiophagus hannah ), which lives throughout South East Asia. This toxin belongs to the cysteine-rich secretory protein (CRISP) family. Ophanin weakly blocks the contraction of smooth muscles elicited by high potassium-induced depolarization, [1] suggesting that it inhibits voltage-dependent calcium channels.
The toxin was named ophanin after the snake whose venom it is derived from, the King Cobra ( Ophiophagus hannah ). [1]
Ophanin is produced in the venom glands of the King Cobra (O. Hannah).
Although the venom has relatively low toxicity, this is compensated by the high amounts of it injected into the prey for each bite. [2]
Ophanin was successfully isolated from O. Hannah venom by gel filtration and cation-exchange chromatography. [1] Its molecular weight is 25 kDa (from positions 19 – 239), which conforms to the molecular mass predicted from its cDNA sequences. [3]
Ophanin is a cysteine-rich secretory protein and therefore belongs to the CRISP family. These proteins possess 16 strictly conserved cysteines and contain 8 disulfide bonds. Ten of the 16 cysteine residues are clustered at the C-terminal end of the protein. Ophanin belongs to the “long” CRISPs subgroup, which consists of the 9 CRISPs with the longest sequences. Snake venom CRISPs belonging to different subgroups act on different biological targets, contributing in this way to the diversity of damaging effects of snake venoms. [4]
The phylogenetic tree constructed from the nucleotide sequences of all known snake venom CRISPs shows that ophanin is more closely related to the Viperidae branch than the Elapidae branch even though O. Hannah belongs to the Elapidae snakes. [1]
Ophanin, along with other specific snake toxins like triflin and ablomin, is also a helothermine-related venom protein (Helveprin) which was originally isolated from the skin of the Mexican beaded lizard. [5]
Ophanin is a weak blocker of the high potassium-induced contraction of smooth muscles. Snake venom CRISP family proteins inhibit depolarization-induced smooth muscle contraction to different extents. Compared to the normal contraction of smooth muscle, ophanin is able to reduce their force of contractility to 84% ± 1%, which is less than most other CRISPs. [1]
The differences between the inhibitory activity of CRISPs may be explained through sequence comparisons that suggest a site that may be critical for inhibition of channel activity. Phe189 and Glu186 are the most likely functional residues: strong blockers of smooth muscle contraction (ablomin, triflin, and latisemin) all have Phe189, and all blockers of smooth muscle contraction except ophanin have Glu186. The significance of this lack of the probable functional residues in ophanin has not yet been addressed. However, it is likely that the picture is more complex and other residues contribute to the inhibitory activity of CRISPs on smooth muscle contraction and some data supports this. For example, pseudecin, while also having Phe189, does not affect depolarization-induced contraction. [1]
There is no direct evidence of a particular mode of action of ophanin blocking depolarization-induced contractions of the smooth muscles. However, based on the hypothesis of Yamazaki and colleagues [6] in regards to ablomin, another snake venom toxin from the CRISP family that also blocks depolarization-induced smooth muscle contraction, we can postulate a similar mechanism might be in place for ophanin.
Since ablomin only blocks contraction induced by depolarization, but not by caffeine, the effect of ablomin is likely to be caused by inhibition of voltage-gated ion channels. An activation of smooth muscle cells through caffeine activates ryanodine receptors of the sarcoplasmic reticulum, whereas an activation through high levels of extracellular potassium depolarizes the membrane (due to the change of the reversal potential for potassium towards more positive values) and would then activate voltage-gated calcium-ion channels leading to high levels of intracellular calcium ions. The intracellular calcium ion concentration correlates well with contraction force in the rat-tail artery. [7] Thus, contraction following extracellular application of high-potassium solution depends on the influx of the extracellular calcium ions through voltage-gated calcium channels. Therefore, ablomin (and by extension ophanin) most likely targets voltage-gated calcium channels on smooth muscle. [6]
The LD50 of the venom in mice is ~1.2 to 3.5 mg/kg via intravenous injection. [8] The LD50 of ophanin is not yet known.
A neuromuscular junction is a chemical synapse between a motor neuron and a muscle fiber.

The cardiac action potential is a brief change in voltage across the cell membrane of heart cells. This is caused by the movement of charged atoms between the inside and outside of the cell, through proteins called ion channels. The cardiac action potential differs from action potentials found in other types of electrically excitable cells, such as nerves. Action potentials also vary within the heart; this is due to the presence of different ion channels in different cells.
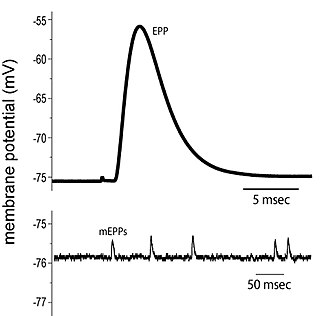
End plate potentials (EPPs) are the voltages which cause depolarization of skeletal muscle fibers caused by neurotransmitters binding to the postsynaptic membrane in the neuromuscular junction. They are called "end plates" because the postsynaptic terminals of muscle fibers have a large, saucer-like appearance. When an action potential reaches the axon terminal of a motor neuron, vesicles carrying neurotransmitters are exocytosed and the contents are released into the neuromuscular junction. These neurotransmitters bind to receptors on the postsynaptic membrane and lead to its depolarization. In the absence of an action potential, acetylcholine vesicles spontaneously leak into the neuromuscular junction and cause very small depolarizations in the postsynaptic membrane. This small response (~0.4mV) is called a miniature end plate potential (MEPP) and is generated by one acetylcholine-containing vesicle. It represents the smallest possible depolarization which can be induced in a muscle.
Voltage-gated calcium channels (VGCCs), also known as voltage-dependent calcium channels (VDCCs), are a group of voltage-gated ion channels found in the membrane of excitable cells (e.g., muscle, glial cells, neurons, etc.) with a permeability to the calcium ion Ca2+. These channels are slightly permeable to sodium ions, so they are also called Ca2+-Na+ channels, but their permeability to calcium is about 1000-fold greater than to sodium under normal physiological conditions.

A conotoxin is one of a group of neurotoxic peptides isolated from the venom of the marine cone snail, genus Conus.
Dendrotoxins are a class of presynaptic neurotoxins produced by mamba snakes (Dendroaspis) that block particular subtypes of voltage-gated potassium channels in neurons, thereby enhancing the release of acetylcholine at neuromuscular junctions. Because of their high potency and selectivity for potassium channels, dendrotoxins have proven to be extremely useful as pharmacological tools for studying the structure and function of these ion channel proteins.

Calciseptine (CaS) is a natural neurotoxin isolated from the black mamba Dendroaspis p. polylepis venom. This toxin consists of 60 amino acids with four disulfide bonds. Calciseptine specifically blocks L-type calcium channels, but not other voltage-dependent Ca2+ channels such as N-type and T-type channels.

Margatoxin (MgTX) is a peptide that selectively inhibits Kv1.3 voltage-dependent potassium channels. It is found in the venom of Centruroides margaritatus, also known as the Central American Bark Scorpion. Margatoxin was first discovered in 1993. It was purified from scorpion venom and its amino acid sequence was determined.
Ablomin is a toxin present in the venom of the Japanese Mamushi snake, which blocks L-type voltage-gated calcium channels.
Triflin is a cysteine-rich secretory protein (CRISP), which is excreted by the venom gland of the Habu snake. Triflin reduces high potassium-induced smooth muscle contraction, suggesting a blocking effect on L-type calcium channels.
Piscivorin is a component of snake venom secreted by the Eastern Cottonmouth. It is a member of the cysteine-rich secretory protein (CRISP) family, which blocks voltage-dependent calcium channels.
Latisemin is a cysteine-rich secretory protein that can be isolated from the venom of the Black-banded sea krait, a sea snake indigenous to the warmer waters of the western Pacific Ocean. It is a toxin that inhibits cyclic nucleotide-gated ion channels and blocks L-type calcium channels, thereby reducing smooth muscle contraction.

Cysteine-rich secretory proteins, often abbreviated as CRISPs, are a group of glycoproteins. They are a subgroup of the CRISP, antigen 5 and Pr-1 (CAP) protein superfamily and also contain a domain related to the ShK toxins. They are substantially implicated in the functioning of the mammalian reproductive system. CRISPs are also found in a variety of snake venoms where they inhibit both smooth muscle contraction and cyclic nucleotide-gated ion channels.
The CAP superfamily is a large superfamily of secreted proteins that are produced by a wide range of organisms, including prokaryotes and non-vertebrate eukaryotes.
Helothermine is a toxin from the venom of the Mexican beaded lizard Heloderma horridum horridum. Helothermine inhibits ryanodine receptors, calcium channels and potassium channels. Helothermine can cause lethargy, partial paralysis of rear limbs and lowering of the body temperature.
Huwentoxins (HWTX) are a group of neurotoxic peptides found in the venom of the Chinese bird spider Haplopelma schmidti. The species was formerly known as Haplopelma huwenum, Ornithoctonus huwena and Selenocosmia huwena. While structural similarity can be found among several of these toxins, HWTX as a group possess high functional diversity.
Tamulotoxin is a venomous neurotoxin from the Indian Red Scorpion.
Pi4 is a short toxin from the scorpion Pandinus imperator that blocks specific potassium channels.

Three-finger toxins are a protein superfamily of small toxin proteins found in the venom of snakes. Three-finger toxins are in turn members of a larger superfamily of three-finger protein domains which includes non-toxic proteins that share a similar protein fold. The group is named for its common structure consisting of three beta strand loops connected to a central core containing four conserved disulfide bonds. The 3FP protein domain has no enzymatic activity and is typically between 60-74 amino acid residues long. Despite their conserved structure, three-finger toxin proteins have a wide range of pharmacological effects. Most members of the family are neurotoxins that act on cholinergic intercellular signaling; the alpha-neurotoxin family interacts with muscle nicotinic acetylcholine receptors (nAChRs), the kappa-bungarotoxin family with neuronal nAChRs, and muscarinic toxins with muscarinic acetylcholine receptors (mAChRs).
OdK2 is a toxin found in the venom of the Iranian scorpion Odonthobuthus doriae. It belongs to the α-KTx family, and selectively blocks the voltage-gated potassium channel Kv1.3 (KCNA3).