Cladistics is an approach to biological classification in which organisms are categorized in groups ("clades") based on hypotheses of most recent common ancestry. The evidence for hypothesized relationships is typically shared derived characteristics (synapomorphies) that are not present in more distant groups and ancestors. However, from an empirical perspective, common ancestors are inferences based on a cladistic hypothesis of relationships of taxa whose character states can be observed. Theoretically, a last common ancestor and all its descendants constitute a (minimal) clade. Importantly, all descendants stay in their overarching ancestral clade. For example, if the terms worms or fishes were used within a strict cladistic framework, these terms would include humans. Many of these terms are normally used paraphyletically, outside of cladistics, e.g. as a 'grade', which are fruitless to precisely delineate, especially when including extinct species. Radiation results in the generation of new subclades by bifurcation, but in practice sexual hybridization may blur very closely related groupings.
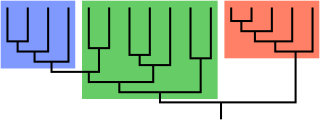
Paraphyly is a taxonomic term describing a grouping that consists of the grouping's last common ancestor and most of its descendants, but excludes one or more subgroups. The grouping is said to be paraphyletic with respect to the excluded subgroups. In contrast, a monophyletic grouping includes a common ancestor and all of its descendants.

Reptiles, as commonly defined, are a group of tetrapods with an ectothermic ('cold-blooded') metabolism and amniotic development. Living reptiles comprise four orders: Testudines (turtles), Crocodilia (crocodilians), Squamata, and Rhynchocephalia. As of May 2023, about 12,000 living species of reptiles are listed in the Reptile Database. The study of the traditional reptile orders, customarily in combination with the study of modern amphibians, is called herpetology.

A cladogram is a diagram used in cladistics to show relations among organisms. A cladogram is not, however, an evolutionary tree because it does not show how ancestors are related to descendants, nor does it show how much they have changed, so many differing evolutionary trees can be consistent with the same cladogram. A cladogram uses lines that branch off in different directions ending at a clade, a group of organisms with a last common ancestor. There are many shapes of cladograms but they all have lines that branch off from other lines. The lines can be traced back to where they branch off. These branching off points represent a hypothetical ancestor which can be inferred to exhibit the traits shared among the terminal taxa above it. This hypothetical ancestor might then provide clues about the order of evolution of various features, adaptation, and other evolutionary narratives about ancestors. Although traditionally such cladograms were generated largely on the basis of morphological characters, DNA and RNA sequencing data and computational phylogenetics are now very commonly used in the generation of cladograms, either on their own or in combination with morphology.

In biology, homology is similarity due to shared ancestry between a pair of structures or genes in different taxa. A common example of homologous structures is the forelimbs of vertebrates, where the wings of bats and birds, the arms of primates, the front flippers of whales, and the forelegs of four-legged vertebrates like dogs and crocodiles are all derived from the same ancestral tetrapod structure. Evolutionary biology explains homologous structures adapted to different purposes as the result of descent with modification from a common ancestor. The term was first applied to biology in a non-evolutionary context by the anatomist Richard Owen in 1843. Homology was later explained by Charles Darwin's theory of evolution in 1859, but had been observed before this, from Aristotle onwards, and it was explicitly analysed by Pierre Belon in 1555.

Amniotes are tetrapod vertebrate animals belonging to the clade Amniota, a large group that comprises the vast majority of living terrestrial and semiaquatic vertebrates. Amniotes evolved from amphibian ancestors during the Carboniferous period and further diverged into two groups, namely the sauropsids and synapsids. They are distinguished from the other living tetrapod clade — the non-amniote lissamphibians — by the development of three extraembryonic membranes, thicker and keratinized skin, and costal respiration.

Sauria is the clade containing the most recent common ancestor of Archosauria and Lepidosauria, and all its descendants. Since most molecular phylogenies recover turtles as more closely related to archosaurs than to lepidosaurs as part of Archelosauria, Sauria can be considered the crown group of diapsids, or reptiles in general. Depending on the systematics, Sauria includes all modern reptiles or most of them as well as various extinct groups.

Sauropsida is a clade of amniotes, broadly equivalent to the class Reptilia, though typically used in a broader sense to include both extinct stem-group relatives of modern reptiles, as well as birds. The most popular definition states that Sauropsida is the sibling taxon to Synapsida, the other clade of amniotes which includes mammals as its only modern representatives. Although early synapsids have historically been referred to as "mammal-like reptiles", all synapsids are more closely related to mammals than to any modern reptile. Sauropsids, on the other hand, include all amniotes more closely related to modern reptiles than to mammals. This includes Aves (birds), which are now recognized as a subgroup of archosaurian reptiles despite originally being named as a separate class in Linnaean taxonomy.

Archosauria is a clade of diapsid sauropsid tetrapods, with birds and crocodilians being the only living representatives. Archosaurs are broadly classified as reptiles, in the cladistic sense of the term, which includes birds. Extinct archosaurs include non-avian dinosaurs, pterosaurs and extinct relatives of crocodilians. Modern paleontologists define Archosauria as a crown group that includes the most recent common ancestor of living birds and crocodilians, and all of its descendants. The base of Archosauria splits into two clades: Pseudosuchia, which includes crocodilians and their extinct relatives; and Avemetatarsalia, which includes birds and their extinct relatives.

In cladistics or phylogenetics, an outgroup is a more distantly related group of organisms that serves as a reference group when determining the evolutionary relationships of the ingroup, the set of organisms under study, and is distinct from sociological outgroups. The outgroup is used as a point of comparison for the ingroup and specifically allows for the phylogeny to be rooted. Because the polarity (direction) of character change can be determined only on a rooted phylogeny, the choice of outgroup is essential for understanding the evolution of traits along a phylogeny.
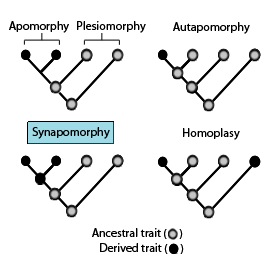
In phylogenetics, an apomorphy is a novel character or character state that has evolved from its ancestral form. A synapomorphy is an apomorphy shared by two or more taxa and is therefore hypothesized to have evolved in their most recent common ancestor. In cladistics, synapomorphy implies homology.

Archosauromorpha is a clade of diapsid reptiles containing all reptiles more closely related to archosaurs rather than lepidosaurs. Archosauromorphs first appeared during the late Middle Permian or Late Permian, though they became much more common and diverse during the Triassic period.
In phylogenetics, a primitive character, trait, or feature of a lineage or taxon is one that is inherited from the common ancestor of a clade and has undergone little change since. Conversely, a trait that appears within the clade group is called advanced or derived. A clade is a group of organisms that consists of a common ancestor and all its lineal descendants.
A grade is a taxon united by a level of morphological or physiological complexity. The term was coined by British biologist Julian Huxley, to contrast with clade, a strictly phylogenetic unit.
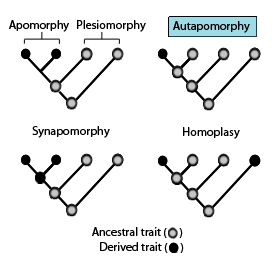
In phylogenetics, an autapomorphy is a distinctive feature, known as a derived trait, that is unique to a given taxon. That is, it is found only in one taxon, but not found in any others or outgroup taxa, not even those most closely related to the focal taxon. It can therefore be considered an apomorphy in relation to a single taxon. The word autapomorphy, introduced in 1950 by German entomologist Willi Hennig, is derived from the Greek words αὐτός, autos "self"; ἀπό, apo "away from"; and μορφή, morphḗ = "shape".

Drepanosaurs are a group of extinct reptiles that lived between the Carnian and Rhaetian stages of the late Triassic Period, approximately between 230 and 210 million years ago. The various species of drepanosaurid were characterized by specialized grasping limbs and often prehensile tails, adaptions for arboreal (tree-dwelling) and fossorial (digging) lifestyles, with some having also been suggested to be aquatic. Fossils of drepanosaurs have been found in Arizona, New Mexico, New Jersey, Utah, England, and northern Italy. The name is taken from the family's namesake genus Drepanosaurus, which means "sickle lizard," a reference to their strongly curved claws.
Phylogenetic nomenclature is a method of nomenclature for taxa in biology that uses phylogenetic definitions for taxon names as explained below. This contrasts with the traditional approach, in which taxon names are defined by a type, which can be a specimen or a taxon of lower rank, and a description in words. Phylogenetic nomenclature is currently regulated by the International Code of Phylogenetic Nomenclature (PhyloCode).

Suchia is a clade of archosaurs containing the majority of pseudosuchians. It was defined as the least inclusive clade containing Aetosaurus ferratus, Rauisuchus tiradentes, Prestosuchus chiniquensis, and Crocodylus niloticus by Nesbitt (2011). Generally the only pseudosuchian group which is omitted from Suchia is the family Ornithosuchidae, although at least one analysis classifies ornithosuchids as close relatives of erpetosuchids and aetosaurs. Phytosaurs are also excluded from Suchia, although it is not certain whether they qualify as pseudosuchians in the first place.
Reptiles arose about 320 million years ago during the Carboniferous period. Reptiles, in the traditional sense of the term, are defined as animals that have scales or scutes, lay land-based hard-shelled eggs, and possess ectothermic metabolisms. So defined, the group is paraphyletic, excluding endothermic animals like birds that are descended from early traditionally-defined reptiles. A definition in accordance with phylogenetic nomenclature, which rejects paraphyletic groups, includes birds while excluding mammals and their synapsid ancestors. So defined, Reptilia is identical to Sauropsida.

Archelosauria is a clade grouping turtles and archosaurs and their fossil relatives, to the exclusion of lepidosaurs. The majority of phylogenetic analyses based on molecular data have supported a sister-group relationship between turtles and archosaurs. On the other hand, Archelosauria had not been historically supported by most morphological analyses, which have instead found turtles to either be descendants of parareptiles, early-diverging diapsids outside of Sauria, or close relatives of lepidosaurs within the clade Ankylopoda. Some recent morphological analyses have also found support for Archelosauria.

