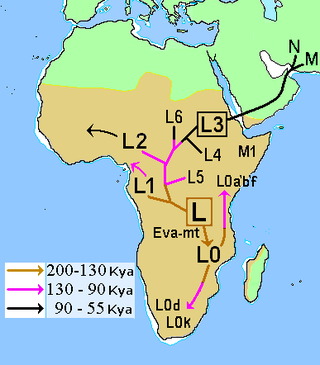
In 200,000 BP, Africans (e.g., Khoisan of Southern Africa) bearing haplogroup L0 diverged from other Africans bearing haplogroup L1′6, which tend to be northward of Southern Africa.[6] Between 130,000 BP and 75,000 BP, behavioral modernity emerged among Southern Africans and long-term interactions between the regions of Southern Africa and Eastern Africa became established.[6]
Among the Klasies River Caves, finger bones (manual distal phalanges), which have been dated between 100,000 BP and 90,000 BP, were recovered from the Witness Baulk in Cave 1.[8] These bones are from an adult individual and appear smaller in size than modern human populations; they are also not comparable to Neanderthal phalanges. These phalanges are however similar to phalanges originating in Die Kelders cave which the authors suggest are more comparable to Holocene Khoesan populations.[8]
In 92,000 BP, amid the Middle Stone Age, Malawian foragers utilized fire to influence and alter their surrounding environment.[9]
At Blombos Cave, small rock fragments with drawn abstractions were "a prime indicator of modern cognition" inscribed by Southern Africans in 73,000 BP.[10] At PP13B, the evidence for symbolic behaviour comes in the form of scraped and ground ochre (usually referred to as limonite bearing powders) that may have been used to form a pigment for body painting. This is similar to more complex ochre utilisation known from Blombos Cave slightly farther to the west at roughly 70,000 years ago.[11] These discoveries contradict the classical hypothesis that the modern behaviour emerged only 40,000 years ago and was reached through a "large cultural leap".[12] The harsh climate and reduced food resources may have been why people moved to the shore at Pinnacle Point, where they could eat marine creatures like shellfish, whale, and seal.[13]
The Later Stone AgeSangoan industry occupied Southern Africa in areas where annual rainfall is less than a meter (1000 mm; 39.4 in).[19]
Various examples of early human technology have been found at Sibudu Cave:
A bone point, a possible arrowhead that pushes back the origin of bow and bone arrow technology to 61,000 BP, at least 20,000 years earlier than the previous earliest example;[20][21]
The earliest known bone needle, dated to 61,000 BP, with wear similar to that found in bone needles used to puncture animal hide;[20]
The earliest example of a compound glue (plant gum and red ochre), used for hafting stone points into wood handles to create spears — dated no later than 71,000 BP;[20] and
Shell beads, although of a more recent date than those found at Blombos cave 71,000 BP for the Sibudu beads in comparison to 75,000 BP for those at Blombos;[22][23]
An example of the use of bedding, dating back to approximately 77,000 BP, with Cape laurel being used on top, probably for its insecticidal properties;[24]
The earliest use of milk (casein) as a paint binder in a milk-ochre mixture 49,000 BP.[25]
Like the earlier Stillbay industry, creators of Howiesons Poort artifacts seem to have engaged in symbolic behavior, having left behind engraved ochre, ostrich eggshells, and shell beads;[27][28] the site of Howiesons Poort dates to at least 50,000 BP.[29] There is a particularly abundant and diverse use of ochre as a pigment for objects or skin, which has been interpreted as reflecting an increasingly complex symbolic culture.[30]
At Border Cave, a nearly complete infant skeleton, which was accompanied by perforated Conus bairstowi shells that dated to ~33,570 ± 120 BP, likely came from the eastern Cape coast.[31] The infant burial may represent one of the earliest southern African examples of burial ornamentation.[32]
At Hora 1 rockshelter, in Malawi, an individual, dated between 16,897 BP and 15,827 BP, carried haplogroups B2b and L5b.[36]
At Hora 1 rockshelter, in Malawi, an individual, dated between 16,424 BP and 14,029 BP, carried haplogroups B2b1a2~ and L0d3/L0d3b.[36]
At Hora, in Malawi, an individual, estimated to date between 10,000 BP and 5000 BP, carried haplogroups BT and L0k2.[37]
Rock art occurs in the form of parietal paintings within the first 40m (130ft) from the entrance of Wonderwerk Cave, possibly all less than 1000 years old, and small engraved stones found within the deposit, mainly from the Later Stone Age sequence where they date back some 10,500 years.[38]
During the early period of the Holocene, in 9000 BP, Khoisan-related peoples admixed with the ancestors of the Igbo people, possibly in the western Sahara.[39][40]
At Hora, in Malawi, an individual, estimated to date between 8173 BP and 7957 BP, carried haplogroup L0a2.[37]
At Fingira rockshelter, in Malawi, an individual, dated between 6179 BP and 2341 BP, carried haplogroups B2 and L0d1.[36]
At Fingira, in Malawi, an individual, estimated to date between 6177 BP and 5923 BP, carried haplogroups BT and L0d1c.[37]
At Fingira, in Malawi, an individual, estimated to date between 6175 BP and 5913 BP, carried haplogroups BT and L0d1b2b.[37]
Bantu-speaking peoples migrated, along with their ceramics, from West Africa into other areas of Sub-Saharan Africa.[43] The Kalundu ceramic type may have spread into Southeastern Africa.[43] Additionally, the Eastern AfricanUrewe ceramic type of Lake Victoria may have spread, via African shores near the Indian Ocean, as the Kwale ceramic type, and spread, via Zimbabwe, Zambia, and Malawi, as the Nkope ceramic type.[43] From the region of Kenya and Tanzania to South Africa, eastern Bantu-speaking Africans constitute a north to south genetic cline; additionally, from eastern Africa to toward southern Africa, evidence of genetic homogeneity is indicative of a serial founder effect and admixture events having occurred between Bantu-speaking Africans and other African populations by the time the Bantu migration had spanned into South Africa.[41] Though some may have been created later, the earlier red finger-painted rock art may have been created between 6000 BP and 1800 BP, to the south of Kei River and Orange River by Khoisan hunter-gatherer-herders, in Malawi and Zambia by considerably dark-skinned, occasionally bearded, bow-and-arrow-wielding Akafula hunter-gatherers who resided in Malawi until 19th century CE, and in Transvaal by the Vhangona people.[44] Bantu-speaking farmers, or their Proto-Bantu progenitors, created the later white finger-painted rock art in some areas of Tanzania, Malawi, Angola, Zambia, and Zimbabwe, as well as in the northern regions of Mozambique, Botswana, and Transvaal.[44] The Transvaal (e.g., Soutpansberg, Waterberg) rock art was specifically created by Sotho-speakers (e.g., Birwa, Koni, Tlokwa) and Venda people.[44] Concentric circles, stylized humans, stylized animals, ox-wagons, saurian figures, Depictions of crocodiles and snakes were included in the white finger-painted rock art tradition, both of which were associated with rainmaking and crocodiles in particular, were also associated with fertility.[44] The white finger-painted rock art may have been created for reasons relating to initiation rites and puberty rituals.[44] Depictions from the rock art tradition of Bantu-speaking farmers have been found on divination-related items (e.g., drums, initiation figurines, initiation masks); fertility terracotta masks from Transvaal have been dated to the 1st millennium CE.[44] Along with Iron Age archaeological sites from the 1st millennium CE, this indicates that white finger-painted rock art tradition may have been spanned from the Early Iron Age to the Later Iron Age.[44] Down-headed animals, which appear in South African rock art, and portray shamanic animal sacrifice as a rainfall ritual, also appear in Round Head rock art.[45]
At Chencherere, in Malawi, an individual, estimated to date between 5400 BP and 4800 BP, carried haplogroup L0k2.[37]
At Chencherere, in Malawi, an individual, estimated to date between 5293 BP and 4979 BP, carried haplogroup L0k1.[37]
Eland Main Panel by Southern San, 4000 BP - 100 BP
At Fingira, in Malawi, an individual, estimated to date between 2676 BP and 2330 BP, carried haplogroup L0f.[37]
As pastoralists and Bantu-speaking agro-pastoralists may have begun arriving in Southern Africa in 2300 BP, Bantu-speaking agropastoralists and Khoisan hunter-gatherers may have admixed with one another, resulting in the development of blended agro-pastoralist and hunter-gatherer communities that languages with click consonants and Khoisanloan words; these amalgamated communities later would develop into the modern indigenous communities (e.g., Xhosa, Sotho, Tswana, Zulu people) of South Africa, Botswana, and Namibia.[49]
At Faraoskop Rock Shelter, in South Africa, an individual, estimated to date between 2017 BP and 1748 BP, carried haplogroups A1b1b2a and L0d1b2b1b.[37]
At Vaalkrans Shelter, in South Africa, an individual, estimated to date to 200 BP, is predominantly related to Khoisan speakers, partly related (15% - 32%) to East Africans, and carried haplogroups L0d3b1.[54]
Related Research Articles
KhoisanKOY-sahn, or Khoe-Sān, is a catch-all term for the indigenous peoples of Southern Africa who traditionally speak non-Bantu languages, combining the Khoekhoen and the Sān peoples. Khoisan populations traditionally speak click languages and are considered to be the historical communities throughout Southern Africa, remaining predominant until European colonisation in areas climatically unfavorable to Bantu (sorghum-based) agriculture, such as the Cape region, through to Namibia, where Khoekhoe populations of Nama and Damara people are prevalent groups, and Botswana. Considerable mingling with Bantu-speaking groups is evidenced by prevalence of click phonemes in many especially Xhosa Southern African Bantu languages.
Behavioral modernity is a suite of behavioral and cognitive traits believed to distinguish current Homo sapiens from other anatomically modern humans, hominins, and primates. Most scholars agree that modern human behavior can be characterized by abstract thinking, planning depth, symbolic behavior, music and dance, exploitation of large game, and blade technology, among others. Underlying these behaviors and technological innovations are cognitive and cultural foundations that have been documented experimentally and ethnographically by evolutionary and cultural anthropologists. These human universal patterns include cumulative cultural adaptation, social norms, language, and extensive help and cooperation beyond close kin.
Africa has the longest record of human habitation in the world. The first hominins emerged 6-7 million years ago, and among the earliest anatomically modern human skulls found so far were discovered at Omo Kibish,Jebel Irhoud, and Florisbad.
Haplogroup A is a human Y-chromosome DNA haplogroup, which includes all living human Y chromosomes. Bearers of extant sub-clades of haplogroup A are almost exclusively found in Africa, in contrast with haplogroup BT, bearers of which participated in the Out of Africa migration of early modern humans. The known branches of haplogroup A are A00, A0, A1a, and A1b1; these branches are only very distantly related, and are not more closely related to each other than they are to haplogroup BT.
Haplogroup E-M96 is a human Y-chromosome DNA haplogroup. It is one of the two main branches of the older and ancestral haplogroup DE, the other main branch being haplogroup D. The E-M96 clade is divided into two main subclades: the more common E-P147, and the less common E-M75.
The Middle Stone Age was a period of African prehistory between the Early Stone Age and the Late Stone Age. It is generally considered to have begun around 280,000 years ago and ended around 50–25,000 years ago. The beginnings of particular MSA stone tools have their origins as far back as 550–500,000 years ago and as such some researchers consider this to be the beginnings of the MSA. The MSA is often mistakenly understood to be synonymous with the Middle Paleolithic of Europe, especially due to their roughly contemporaneous time span; however, the Middle Paleolithic of Europe represents an entirely different hominin population, Homo neanderthalensis, than the MSA of Africa, which did not have Neanderthal populations. Additionally, current archaeological research in Africa has yielded much evidence to suggest that modern human behavior and cognition was beginning to develop much earlier in Africa during the MSA than it was in Europe during the Middle Paleolithic. The MSA is associated with both anatomically modern humans as well as archaic Homo sapiens, sometimes referred to as Homo helmei. Early physical evidence comes from the Gademotta Formation in Ethiopia, the Kapthurin Formation in Kenya and Kathu Pan in South Africa.
The Prehistory of South Africa lasts from the Middle Stone Age until the 17th century. Southern Africa was first reached by Homo sapiens before 130,000 years ago, possibly before 260,000 years ago. The region remained in the Late Stone Age until the first traces of pastoralism were introduced about 2,000 years ago. The Bantu migration reached the area now South Africa around the first decade of the 3rd century, over 1800 years ago. Early Bantu kingdoms were established in the 11th century. First European contact dates to 1488, but European colonization began in the 17th century.
Sibudu Cave is a rock shelter in a sandstone cliff in northern KwaZulu-Natal, South Africa. It is an important Middle Stone Age site occupied, with some gaps, from 77000 years ago to 38000 years ago.
In human mitochondrial genetics, L is the mitochondrial DNA macro-haplogroup that is at the root of the anatomically modern human mtDNA phylogenetic tree. As such, it represents the most ancestral mitochondrial lineage of all currently living modern humans, also dubbed "Mitochondrial Eve".
Border Cave is an archaeological site located in the western Lebombo Mountains in Kwazulu-Natal. The rock shelter has one of the longest archaeological records in southern Africa, which spans from the Middle Stone Age to the Iron Age.
Haplogroup E-M2, also known as E1b1a1-M2, is a human Y-chromosome DNA haplogroup. E-M2 is primarily distributed within Africa followed by West Asia. More specifically, E-M2 is the predominant subclade in West Africa, Central Africa, Southern Africa, and the region of the African Great Lakes; it also occurs at moderate frequencies in North Africa, and the Middle East. E-M2 has several subclades, but many of these subhaplogroups are included in either E-L485 or E-U175. E-M2 is especially common among indigenous Africans who speak Niger-Congo languages, and was spread to Southern Africa and East Africa through the Bantu expansion.
The genetic history of Africa summarizes the genetic makeup and population history of African populations in Africa, composed of the overall genetic history, including the regional genetic histories of North Africa, West Africa, East Africa, Central Africa, and Southern Africa, as well as the recent origin of modern humans in Africa. The Sahara served as a trans-regional passageway and place of dwelling for people in Africa during various humid phases and periods throughout the history of Africa.
The history of East Africa has been divided into its prehistory, the major polities flourishing, the colonial period, and the post-colonial period, in which the current nations were formed. East Africa is the eastern region of Africa, bordered by North Africa, Central Africa, Southern Africa, the Indian Ocean, and the Sahara Desert. Colonial boundaries are reflected in the modern boundaries between contemporary East African states, cutting across ethnic and cultural lines, often dividing single ethnic groups between two or more states.
The history of Southern Africa has been divided into its prehistory, its ancient history, the major polities flourishing, the colonial period, and the post-colonial period, in which the current nations were formed. Southern Africa is bordered by Central Africa, East Africa, the Atlantic Ocean, the Indian Ocean, and the Sahara Desert. Colonial boundaries are reflected in the modern boundaries between contemporary Southern African states, cutting across ethnic and cultural lines, often dividing single ethnic groups between two or more states.
The genetic history of the African diaspora is composed of the overall genetic history of the African diaspora, within regions outside of Africa, such as North America, Central America, the Caribbean, South America, Europe, Asia, and Australia; this includes the genetic histories of African Americans, Afro-Canadians, Afro-Caribbeans, Afro-Latinos, Afro-Europeans, Afro-Asians, and African Australians.
The prehistory of East Africa spans from the earliest human presence in the region until the emergence of the Iron Age in East Africa. Between 1,600,000 BP and 1,500,000 BP, the Homo ergaster known as Nariokotome Boy resided near Nariokotome River, Kenya. Modern humans, who left behind remains, resided at Omo Kibish in 233,000 BP. Afro-Asiatic speakers and Nilo-Saharan speakers expanded in East Africa, resulting in transformation of food systems of East Africa. Prehistoric West Africans may have diverged into distinct ancestral groups of modern West Africans and Bantu-speaking peoples in Cameroon, and, subsequently, around 5000 BP, the Bantu-speaking peoples migrated into other parts of Sub-Saharan Africa.
The genetic history of West Africa encompasses the genetic history of the people of West Africa. The Sahara served as a trans-regional passageway and place of dwelling for people in Africa during various humid phases and periods throughout the history of Africa.
The genetic history of Central Africa encompasses the genetic history of the people of Central Africa. The Sahara served as a trans-regional passageway and place of dwelling for people in Africa during various humid phases and periods throughout the history of Africa.
The genetic history of Eastern Africa encompasses the genetic history of the people of Eastern Africa. The Sahara served as a trans-regional passageway and place of dwelling for people in Africa during various humid phases and periods throughout the history of Africa.
The genetic history of Southern Africa encompasses the genetic history of the people of Southern Africa. The Sahara served as a trans-regional passageway and place of dwelling for people in Africa during various humid phases and periods throughout the history of Africa.
1 2 Estimated split times given in the source cited (in kya): Human-Neanderthal: 530-690, Deep Human [H. sapiens]: 250-360, NKSP-SKSP: 150-190, Out of Africa (OOA): 70–120.
↑ Lee, Richard B. (1976), Kalahari Hunter-Gatherers: Studies of the ǃKung San and Their Neighbors, Richard B. Lee and Irven DeVore, eds. Cambridge: Harvard University Press
1 2 3 Backwell, L; d'Errico, F; Wadley, L (2008). "Middle Stone Age bone tools from the Howiesons Poort layers, Sibudu Cave, South Africa". Journal of Archaeological Science. 35 (6): 1566–1580. doi:10.1016/j.jas.2007.11.006.
↑ d'Errico, F; Vanhaeren, M; Wadley, L (2008). "Possible shell beads from the Middle Stone Age layers of Sibudu Cave, South Africa". Journal of Archaeological Science. 35 (10): 2675–2685. Bibcode:2008JArSc..35.2675D. doi:10.1016/j.jas.2008.04.023.
↑ d'Errico, F; Henshilwood, C; Vanhaeren, M; van Niekerk, K (January 2005). "Nassarius kraussianus shell beads from Blombos Cave: evidence for symbolic behaviour in the Middle Stone Age". J. Hum. Evol. 48 (1): 3–24. doi:10.1016/j.jhevol.2004.09.002. PMID15656934.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.