Related Research Articles

Jet lag, desynchronosis, or circadian dysrhythmia, is a temporary physiological condition that occurs when a person's circadian rhythm is out of sync with the time zone they are in, and is a typical result from travelling rapidly across multiple time zones. For example, someone travelling from New York to London, i.e. from west to east, feels as if the time were five hours earlier than local time, and someone travelling from London to New York, i.e. from east to west, feels as if the time were five hours later than local time. The phase shift when travelling from east to west is referred to as phase-delay of the circadian cycle, whereas going west to east is phase-advance of the cycle. Most travellers find that it is harder to adjust time zones when travelling east. Jet lag was previously classified as a circadian rhythm sleep disorder.

A circadian rhythm, or circadian cycle, is a natural oscillation that repeats roughly every 24 hours. Circadian rhythms can refer to any process that originates within an organism and responds to the environment. Circadian rhythms are regulated by a circadian clock whose primary function is to rhythmically co-ordinate biological processes so they occur at the correct time to maximize the fitness of an individual. Circadian rhythms have been widely observed in animals, plants, fungi and cyanobacteria and there is evidence that they evolved independently in each of these kingdoms of life.

Bee learning and communication includes cognitive and sensory processes in all kinds of bees, that is the insects in the seven families making up the clade Anthophila. Some species have been studied more extensively than others, in particular Apis mellifera, or European honey bee. Color learning has also been studied in bumblebees.

Foraging is searching for wild food resources. It affects an animal's fitness because it plays an important role in an animal's ability to survive and reproduce. Foraging theory is a branch of behavioral ecology that studies the foraging behavior of animals in response to the environment where the animal lives.

In cognitive psychology and neuroscience, spatial memory is a form of memory responsible for the recording and recovery of information needed to plan a course to a location and to recall the location of an object or the occurrence of an event. Spatial memory is necessary for orientation in space. Spatial memory can also be divided into egocentric and allocentric spatial memory. A person's spatial memory is required to navigate in a familiar city. A rat's spatial memory is needed to learn the location of food at the end of a maze. In both humans and animals, spatial memories are summarized as a cognitive map.
Waggle dance is a term used in beekeeping and ethology for a particular figure-eight dance of the honey bee. By performing this dance, successful foragers can share information about the direction and distance to patches of flowers yielding nectar and pollen, to water sources, or to new nest-site locations with other members of the colony.

Diurnality is a form of plant and animal behavior characterized by activity during daytime, with a period of sleeping or other inactivity at night. The common adjective used for daytime activity is "diurnal". The timing of activity by an animal depends on a variety of environmental factors such as the temperature, the ability to gather food by sight, the risk of predation, and the time of year. Diurnality is a cycle of activity within a 24-hour period; cyclic activities called circadian rhythms are endogenous cycles not dependent on external cues or environmental factors except for a zeitgeber. Animals active during twilight are crepuscular, those active during the night are nocturnal and animals active at sporadic times during both night and day are cathemeral.

Latent learning is the subconscious retention of information without reinforcement or motivation. In latent learning, one changes behavior only when there is sufficient motivation later than when they subconsciously retained the information.
In psychology and neuroscience, time perception or chronoception is the subjective experience, or sense, of time, which is measured by someone's own perception of the duration of the indefinite and unfolding of events. The perceived time interval between two successive events is referred to as perceived duration. Though directly experiencing or understanding another person's perception of time is not possible, perception can be objectively studied and inferred through a number of scientific experiments. Some temporal illusions help to expose the underlying neural mechanisms of time perception.
A chronotype is the behavioral manifestation of underlying circadian rhythm's myriad of physical processes. A person's chronotype is the propensity for the individual to sleep at a particular time during a 24-hour period. Eveningness and morningness are the two extremes with most individuals having some flexibility in the timing of their sleep period. However, across development there are changes in the propensity of the sleep period with pre-pubescent children preferring an advanced sleep period, adolescents preferring a delayed sleep period and many elderly preferring an advanced sleep period.
Animal psychopathology is the study of mental or behavioral disorders in non-human animals.
Light effects on circadian rhythm are the response of circadian rhythms to light.

Euhirudinea, the true leeches, are an infraclass of the Hirudinea.
Task allocation and partitioning is the way that tasks are chosen, assigned, subdivided, and coordinated within a colony of social insects. Task allocation and partitioning gives rise to the division of labor often observed in social insect colonies, whereby individuals specialize on different tasks within the colony. Communication is closely related to the ability to allocate tasks among individuals within a group. This entry focuses exclusively on social insects. For information on human task allocation and partitioning, see division of labour, task analysis, and workflow.
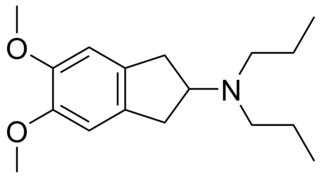
PNU-99,194(A) (or U-99,194(A)) is a drug which acts as a moderately selective D3 receptor antagonist with ~15-30-fold preference for D3 over the D2 subtype. Though it has substantially greater preference for D3 over D2, the latter receptor does still play some role in its effects, as evidenced by the fact that PNU-99,194 weakly stimulates both prolactin secretion and striatal dopamine synthesis, actions it does not share with the more selective (100-fold) D3 receptor antagonists S-14,297 and GR-103,691.
Episodic-like memory is the memory system in animals that is comparable to human episodic memory. The term was first described by Clayton & Dickinson referring to an animal's ability to encode and retrieve information about 'what' occurred during an episode, 'where' the episode took place, and 'when' the episode happened. This ability in animals is considered 'episodic-like' because there is currently no way of knowing whether or not this form of remembering is accompanied by conscious recollection—a key component of Endel Tulving's original definition of episodic memory.
Social learning refers to learning that is facilitated by observation of, or interaction with, another animal or its products. Social learning has been observed in a variety of animal taxa, such as insects, fish, birds, reptiles, amphibians and mammals.
Cognitive bias in animals is a pattern of deviation in judgment, whereby inferences about other animals and situations may be affected by irrelevant information or emotional states. It is sometimes said that animals create their own "subjective social reality" from their perception of the input. In humans, for example, an optimistic or pessimistic bias might affect one's answer to the question "Is the glass half empty or half full?"

Insect cognition describes the mental capacities and study of those capacities in insects. The field developed from comparative psychology where early studies focused more on animal behavior. Researchers have examined insect cognition in bees, fruit flies, and wasps.
The food-entrainable oscillator (FEO) is a circadian clock that can be entrained by varying the time of food presentation. It was discovered when a rhythm was found in rat activity. This was called food anticipatory activity (FAA), and this is when the wheel-running activity of mice decreases after feeding, and then rapidly increases in the hours leading up to feeding. FAA appears to be present in non-mammals (pigeons/fish), but research heavily focuses on its presence in mammals. This rhythmic activity does not require the suprachiasmatic nucleus (SCN), the central circadian oscillator in mammals, implying the existence of an oscillator, the FEO, outside of the SCN, but the mechanism and location of the FEO is not yet known. There is ongoing research to investigate if the FEO is the only non-light entrainable oscillator in the body.
References
- ↑ Mulder, C.K.; Gerkema, M.P.; Van der Zee, E.A. (2013). "Circadian clocks and memory: Time-place learning". Frontiers in Molecular Neuroscience. 6: 1–10. doi: 10.3389/fnmol.2013.00008 . PMC 3622895 . PMID 23596390.
- ↑ Wahl, O (1932). "Neue Untersuchungen über das Zeitgedachtnis der Bienen". Zeitschrift für vergleichende Physiologie. 16 (3): 529–589. doi:10.1007/BF00338333. S2CID 6662886.
- ↑ Moore, D.; Siegfried, D.; Wilson, R.; Rankin, M.A. (1989). "The influence of time of day on the foraging behavior of the honeybee, Apis mellifera". Journal of Biological Rhythms. 4 (3): 305–325. doi: 10.1177/074873048900400301 . PMID 2519596. S2CID 24854091.
- ↑ Zhang, S.; Schwarz, S.; Pahl, M.; Zhu, H.; Tautz, J. (2006). "Honeybee memory: a honeybee knows what to do and when". Journal of Experimental Biology. 209 (22): 4420–4428. doi: 10.1242/jeb.02522 . PMID 17079712.
- ↑ Schatz, B.; Lachaud, J.-P.; Beugnon, G. (1999). "Spatio-temporal learning by the ant Ectatomma ruidum" (PDF). Journal of Experimental Biology. 202 (14): 1897–1907. doi:10.1242/jeb.202.14.1897. PMID 10377271.
- ↑ Reebs, S.G. (1996). "Time-place learning in golden shiners (Pisces: Notemigonus crysoleucas)". Behavioural Processes. 36 (3): 253–262. doi:10.1016/0376-6357(96)88023-5. PMID 24896874. S2CID 12061959.
- ↑ Reebs, S.G. (1999). "Time-place learning based on food but not on predation risk in a fish, the inanga (Galaxias maculatus)". Ethology. 105 (4): 361–371. doi:10.1046/j.1439-0310.1999.00390.x.
- ↑ Reebs, S.G. (1993). "A test of time-place learning in a cichlid fish". Behavioural Processes. 30 (3): 273–282. doi:10.1016/0376-6357(93)90139-I. PMID 24896951. S2CID 20397135.
- ↑ Biebach, H.; Gordijn, M.; Krebs, J.R. (1989). "Time-and-place learning by garden warblers, Sylvia borin". Animal Behaviour. 37: 353–360. doi:10.1016/0003-3472(89)90083-3. S2CID 54382951.
- ↑ Krebs, J.R.; Biebach, H. (1989). "Time-place learning by garden warblers (Sylvia borin): route or map?". Ethology. 83 (3): 248–256. doi:10.1111/j.1439-0310.1989.tb00532.x.
- ↑ Biebach, H.; Falk, H.; Krebs, J.R. (1991). "The effect of constant light and phase-shifts on a learned time-place association in garden warblers (Sylvia borin): hourglass or circadian clock?". Journal of Biological Rhythms. 6 (4): 353–365. doi: 10.1177/074873049100600406 . PMID 1773101. S2CID 36202996.
- ↑ Wenger, D.; Biebach, H.; Krebs, J.R. (1991). "Free-running circadian rhythm of a learned feeding pattern in starlings". Naturwissenschaften. 78 (2): 87–89. Bibcode:1991NW.....78...87W. doi:10.1007/bf01206266. S2CID 34798321.
- ↑ Falk, H.; Biebach, H.; Krebs, J.R. (1992). "Learning a time-place pattern of food availability: a comparison between an insectivorous and a granivorous weaver species (Ploceus bicolor and Euplectes hordeaceus)". Behavioral Ecology and Sociobiology. 31: 9–15. doi:10.1007/bf00167811. S2CID 23105670.
- ↑ Saksida, L.M.; Wilkie, D.M. (1994). "Time-of-day discrimination by pigeons, Columbia livia". Animal Learning & Behavior. 22 (2): 143–154. doi: 10.3758/bf03199914 .
- ↑ Means, L.W.; Ginn, S.R.; Arolfo, M.P.; Pence, J.D. (2000). "Breakfast in the nook and dinner in the dining room: time-of-day discrimination in rats". Behavioural Processes. 49 (1): 21–33. doi:10.1016/s0376-6357(00)00068-1. PMID 10725650. S2CID 35474894.
- ↑ Lukoyanov, N.V.; Pereira, P.A.; Mesquita, R.M.; Andrade, J.P. (2002). "Restricted feeding facilitates time-place learning in adult rats". Behavioural Brain Research. 134 (1–2): 283–290. doi:10.1016/s0166-4328(02)00036-0. PMID 12191815. S2CID 21591389.
- ↑ Widman, D.R.; Sermania, C.M.; Genismore, K.E. (2004). "Evidence for time-place learning in the Morris water maze without food restriction but with increased response cost". Behavioural Processes. 67 (2): 183–193. doi:10.1016/j.beproc.2004.04.001. PMID 15240056. S2CID 11651650.
- ↑ Deibel, S.H.; Thorpe, C.M. (2013). "The effects of response cost and species-typical behaviors on a daily time-place task". Learning & Behavior. 41 (1): 42–53. doi: 10.3758/s13420-012-0076-4 . PMID 22669816.
- ↑ Van der Zee, E.A.; Havekes, R.; Barf, R.R.; Hut, R.A.; Nijholt, I.M.; Jacobs, E.H.; Gerkema, M.P. (2008). "Circadian time-place learning in mice depends on Cry genes". Current Biology. 18 (11): 844–848. doi: 10.1016/j.cub.2008.04.077 . PMID 18514517.
- ↑ Mulder, C.K.; Papantoniou, C.; Gerkema, M.P.; Van der Zee, E.A. (2014). "Neither the SCN nor the adrenals are required for circadian time-place learning in mice". Chronobiology International. 31 (9): 1075–1092. doi:10.3109/07420528.2014.944975. PMC 4219850 . PMID 25083974.