Amniotes are tetrapod vertebrate animals belonging to the clade Amniota, a large group that comprises the vast majority of living terrestrial and semiaquatic vertebrates. Amniotes evolved from amphibian ancestors during the Carboniferous period and further diverged into two groups, namely the sauropsids and synapsids. They are distinguished from the other living tetrapod clade — the non-amniote lissamphibians — by the development of three extraembryonic membranes, thicker and keratinized skin, and costal respiration.

"Labyrinthodontia" is an informal grouping of extinct predatory amphibians which were major components of ecosystems in the late Paleozoic and early Mesozoic eras. Traditionally considered a subclass of the class Amphibia, modern classification systems recognize that labyrinthodonts are not a formal natural group (clade) exclusive of other tetrapods. Instead, they consistute an evolutionary grade, ancestral to living tetrapods such as lissamphibians and amniotes. "Labyrinthodont"-grade vertebrates evolved from lobe-finned fishes in the Devonian, though a formal boundary between fish and amphibian is difficult to define at this point in time.

Lepospondyli is a diverse taxon of early tetrapods. With the exception of one late-surviving lepospondyl from the Late Permian of Morocco, lepospondyls lived from the Early Carboniferous (Mississippian) to the Early Permian and were geographically restricted to what is now Europe and North America. Five major groups of lepospondyls are known: Adelospondyli; Aïstopoda; Lysorophia; Microsauria; and Nectridea. Lepospondyls have a diverse range of body forms and include species with newt-like, eel- or snake-like, and lizard-like forms. Various species were aquatic, semiaquatic, or terrestrial. None were large, and they are assumed to have lived in specialized ecological niches not taken by the more numerous temnospondyl amphibians that coexisted with them in the Paleozoic. Lepospondyli was named in 1888 by Karl Alfred von Zittel, who coined the name to include some tetrapods from the Paleozoic that shared some specific characteristics in the notochord and teeth. Lepospondyls have sometimes been considered to be either related or ancestral to modern amphibians or to Amniota. It has been suggested that the grouping is polyphyletic, with aïstopods being primitive stem-tetrapods, while recumbirostran microsaurs are primitive reptiles.

Adelospondyli is an order of elongated, presumably aquatic, Carboniferous amphibians. They have a robust skull roofed with solid bone, and orbits located towards the front of the skull. The limbs were almost certainly absent, although some historical sources reported them to be present. Despite the likely absence of limbs, adelospondyls retained a large part of the bony shoulder girdle. Adelospondyls have been assigned to a variety of groups in the past. They have traditionally been seen as members of the subclass Lepospondyli, related to other unusual early tetrapods such as "microsaurs", "nectrideans", and aïstopods. Analyses such as Ruta & Coates (2007) have offered an alternate classification scheme, arguing that adelospondyls were actually far removed from other lepospondyls, instead being stem-tetrapod stegocephalians closely related to the family Colosteidae.

Reptiliomorpha is a clade containing the amniotes and those tetrapods that share a more recent common ancestor with amniotes than with living amphibians (lissamphibians). It was defined by Michel Laurin (2001) and Vallin and Laurin (2004) as the largest clade that includes Homo sapiens, but not Ascaphus truei. Laurin and Reisz (2020) defined Pan-Amniota as the largest total clade containing Homo sapiens, but not Pipa pipa, Caecilia tentaculata, and Siren lacertina.

Diadectes is an extinct genus of large reptiliomorphs or synapsids that lived during the early Permian period. Diadectes was one of the first herbivorous tetrapods, and also one of the first fully terrestrial vertebrates to attain large size.
Diadectomorpha is a clade of large tetrapods that lived in Euramerica during the Carboniferous and Early Permian periods and in Asia during Late Permian (Wuchiapingian), They have typically been classified as advanced reptiliomorphs positioned close to, but outside of the clade Amniota, though some recent research has recovered them as the sister group to the traditional Synapsida within Amniota, based on inner ear anatomy and cladistic analyses. They include both large carnivorous and even larger herbivorous forms, some semi-aquatic and others fully terrestrial. The diadectomorphs seem to have originated during late Mississippian times, although they only became common after the Carboniferous rainforest collapse and flourished during the Late Pennsylvanian and Early Permian periods.

Seymouria is an extinct genus of seymouriamorph from the Early Permian of North America and Europe. Although they were amphibians, Seymouria were well-adapted to life on land, with many reptilian features—so many, in fact, that Seymouria was first thought to be a primitive reptile. It is primarily known from two species, Seymouria baylorensis and Seymouria sanjuanensis. The type species, S. baylorensis, is more robust and specialized, though its fossils have only been found in Texas. On the other hand, Seymouria sanjuanensis is more abundant and widespread. This smaller species is known from multiple well-preserved fossils, including a block of six skeletons found in the Cutler Formation of New Mexico, and a pair of fully grown skeletons from the Tambach Formation of Germany, which were fossilized lying next to each other.
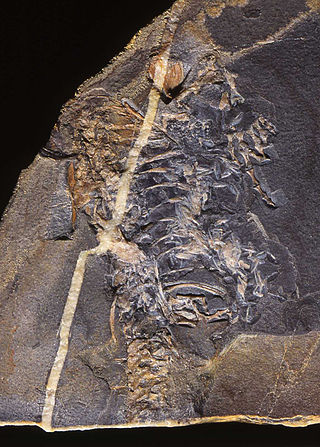
Casineria is an extinct genus of tetrapod which lived about 340-334 million years ago in the Mississippian epoch of the Carboniferous period. Its generic name, Casineria, is a latinization of Cheese Bay. The site near Edinburgh, Scotland where the holotype fossil was found. When originally described in 1999, it was identified as a transitional fossil noted for its mix of basal (amphibian-like) and advanced (reptile-like) characteristics, putting it at or very near the origin of the amniotes, the group containing all mammals, birds, modern reptiles, and other descendants of their reptile-like common ancestor. However, the sole known fossil is lacking key elements such as a skull, making exact analysis difficult. As a result, the classification of Casineria has been more controversial in analyses conducted since 1999. Other proposed affinities include a placement among the lepospondyls, seymouriamorphs, "gephyrostegids", or as a synonym of Caerorhachis, another controversial tetrapod which may have been an early temnospondyl.

Eucritta is an extinct genus of stem-tetrapod from the Viséan epoch in the Carboniferous period of Scotland. The name of the type and only species, E. melanolimnetes is a homage to the 1954 horror film Creature from the Black Lagoon.

Askeptosaurus is an extinct genus of askeptosauroid, a marine reptile from the extinct order Thalattosauria. Askeptosaurus is known from several well-preserved fossils found in Middle Triassic marine strata in what is now Italy and Switzerland.

Embolomeri is an order of tetrapods or stem-tetrapods, possibly members of Reptiliomorpha. Embolomeres first evolved in the Early Carboniferous (Mississippian) Period and were the largest and most successful predatory tetrapods of the Late Carboniferous (Pennsylvanian) Period. They were specialized semiaquatic predators with long bodies for eel-like undulatory swimming. Embolomeres are characterized by their vertebral centra, which are formed by two cylindrical segments, the pleurocentrum at the rear and intercentrum at the front. These segments are equal in size. Most other tetrapods have pleurocentra and intercentra which are drastically different in size and shape.

Limnoscelis was a genus of large diadectomorph tetrapods from the Late Carboniferous to early Permian of western North America. It includes two species: the type species Limnoscelis paludis from New Mexico, and Limnoscelis dynatis from Colorado, both of which are thought to have lived concurrently. No specimens of Limnoscelis are known from outside of North America. Limnoscelis was carnivorous, and likely semiaquatic, though it may have spent a significant portion of its life on land. Limnoscelis had a combination of derived amphibian and primitive reptilian features, and its placement relative to Amniota has significant implications regarding the origins of the first amniotes.

Colosteidae is a family of stegocephalians that lived in the Carboniferous period. They possessed a variety of characteristics from different tetrapod or stem-tetrapod groups, which made them historically difficult to classify. They are now considered to be part of a lineage intermediate between the earliest Devonian terrestrial vertebrates, and the different groups ancestral to all modern tetrapods, such as temnospondyls and reptiliomorphs.

Acherontiscus is an extinct genus of stegocephalians that lived in the Early Carboniferous of Scotland. The type and only species is Acherontiscus caledoniae, named by paleontologist Robert Carroll in 1969. Members of this genus have an unusual combination of features which makes their placement within amphibian-grade tetrapods uncertain. They possess multi-bone vertebrae similar to those of embolomeres, but also a skull similar to lepospondyls. The only known specimen of Acherontiscus possessed an elongated body similar to that of a snake or eel. No limbs were preserved, and evidence for their presence in close relatives of Acherontiscus is dubious at best. Phylogenetic analyses created by Marcello Ruta and other paleontologists in the 2000s indicate that Acherontiscus is part of Adelospondyli, closely related to other snake-like animals such as Adelogyrinus and Dolichopareias. Adelospondyls are traditionally placed within the group Lepospondyli due to their fused vertebrae. Some analyses published since 2007 have argued that adelospondyls such as Acherontiscus may not actually be lepospondyls, instead being close relatives or members of the family Colosteidae. This would indicate that they evolved prior to the split between the tetrapod lineage that leads to reptiles (Reptiliomorpha) and the one that leads to modern amphibians (Batrachomorpha). Members of this genus were probably aquatic animals that were able to swim using snake-like movements.

Odonterpeton is an extinct genus of "microsaur" from the Late Carboniferous of Ohio, containing the lone species Odonterpeton triangulare. It is known from a single partial skeleton preserving the skull, forelimbs, and the front part of the torso. The specimen was found in the abandoned Diamond Coal Mine of Linton, Ohio, a fossiliferous coal deposit dated to the late Moscovian stage, about 310 million years ago.
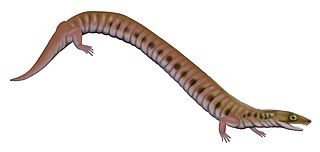
Rhynchonkos is an extinct genus of rhynchonkid microsaur. Originally known as Goniorhynchus, it was renamed in 1981 because the name had already been given to another genus; the family, likewise, was originally named Goniorhynchidae but renamed in 1988. The type and only known species is R. stovalli, found from the Early Permian Fairmont Shale in Cleveland County, Oklahoma. Rhynchonkos shares many similarities with Eocaecilia, an early caecilian from the Early Jurassic of Arizona. Similarities between Rhynchonkos and Eocaecilia have been taken as evidence that caecilians are descendants of microsaurs. However, such a relationship is no longer widely accepted.

Tuditanidae is an extinct family of microsaurian tetrapods. Fossils have been found from Nova Scotia, Ohio, and the Czech Republic and are Late Carboniferous in age.

Microleter is an extinct genus of basal procolophonomorph parareptiles which lived in Oklahoma during the Early Permian period. The type and only known species is Microleter mckinzieorum. Microleter is one of several parareptile taxa described from the Richards Spur fissure fills, and can be characterized from its high tooth count, lacrimal/narial contact, short postfrontal, and slit-like temporal emargination edged by the postorbital, jugal, squamosal, and quadratojugal. Contrary to Australothyris, which had a similar phylogenetic position as a basal procolophonomorph, Microleter suggests that early parareptile evolution occurred in Laurasia and that multiple lineages developed openings or emarginations in the temporal region.

Erpetonyx is an extinct genus of bolosaurian parareptile from the Gzhelian stage of the Carboniferous period, with a single known species: Erpetonyx arsenaultorum. It is known from a single articulated and mostly complete specimen from Prince Edward Island in Canada. Phylogenetics has predicted that parareptiles first evolved in the Carboniferous, parallel to eureptiles. However, Hylonomus, the oldest eureptile known from fossil evidence, lived millions of years before parareptiles appeared in the fossil record. The discovery of Erpetonyx helped to shorten this gap between parareptile and eureptile fossils, as Erpetonyx lived in the Late Carboniferous and is one of the oldest known parareptiles. However, it was not closely related to ancestral parareptiles, so its discovery also indicated that the initial diversification of parareptiles occurred earlier in the Carboniferous. Erpetonyx was a small reptile, with the entire skeleton about 20 to 25 centimeters in length. It was likely carnivorous, and could be characterized by a variety of skeletal features, including a relatively elongated body and large claws with powerful tendon attachment points.