
Dimetrodon is an extinct genus of non-mammalian synapsid belonging to the family Sphenacodontidae that lived during the Cisuralian age of the Early Permian period, around 295–272 million years ago. With most species measuring 1.7–4.6 m (5.6–15.1 ft) long and weighing 28–250 kg (62–551 lb), the most prominent feature of Dimetrodon is the large neural spine sail on its back formed by elongated spines extending from the vertebrae. It was an obligate quadruped and had a tall, curved skull with large teeth of different sizes set along the jaws. Most fossils have been found in the Southwestern United States, the majority of these coming from a geological deposit called the Red Beds of Texas and Oklahoma. More recently, its fossils have also been found in Germany and over a dozen species have been named since the genus was first erected in 1878.

"Labyrinthodontia" is an informal grouping of extinct predatory amphibians which were major components of ecosystems in the late Paleozoic and early Mesozoic eras. Traditionally considered a subclass of the class Amphibia, modern classification systems recognize that labyrinthodonts are not a formal natural group (clade) exclusive of other tetrapods. Instead, they consistute an evolutionary grade, ancestral to living tetrapods such as lissamphibians and amniotes. "Labyrinthodont"-grade vertebrates evolved from lobe-finned fishes in the Devonian, though a formal boundary between fish and amphibian is difficult to define at this point in time.

Reptiliomorpha is a clade containing the amniotes and those tetrapods that share a more recent common ancestor with amniotes than with living amphibians (lissamphibians). It was defined by Michel Laurin (2001) and Vallin and Laurin (2004) as the largest clade that includes Homo sapiens, but not Ascaphus truei. Laurin and Reisz (2020) defined Pan-Amniota as the largest total clade containing Homo sapiens, but not Pipa pipa, Caecilia tentaculata, and Siren lacertina.

Diadectes is an extinct genus of large reptiliomorphs or synapsids that lived during the early Permian period. Diadectes was one of the first herbivorous tetrapods, and also one of the first fully terrestrial vertebrates to attain large size.
Diadectidae is an extinct family of early tetrapods that lived in what is now North America and Europe during the Late Carboniferous and Early Permian, and in Asia during the Late Permian. They were the first herbivorous tetrapods, and also the first fully terrestrial animals to attain large sizes. Footprints indicate that diadectids walked with an erect posture. They were the first to exploit plant material in terrestrial food chains, making their appearance an important stage in both vertebrate evolution and the development of terrestrial ecosystems.
Diadectomorpha is a clade of large tetrapods that lived in Euramerica during the Carboniferous and Early Permian periods and in Asia during Late Permian (Wuchiapingian), They have typically been classified as advanced reptiliomorphs positioned close to, but outside of the clade Amniota, though some recent research has recovered them as the sister group to the traditional Synapsida within Amniota, based on inner ear anatomy and cladistic analyses. They include both large carnivorous and even larger herbivorous forms, some semi-aquatic and others fully terrestrial. The diadectomorphs seem to have originated during late Mississippian times, although they only became common after the Carboniferous rainforest collapse and flourished during the Late Pennsylvanian and Early Permian periods.

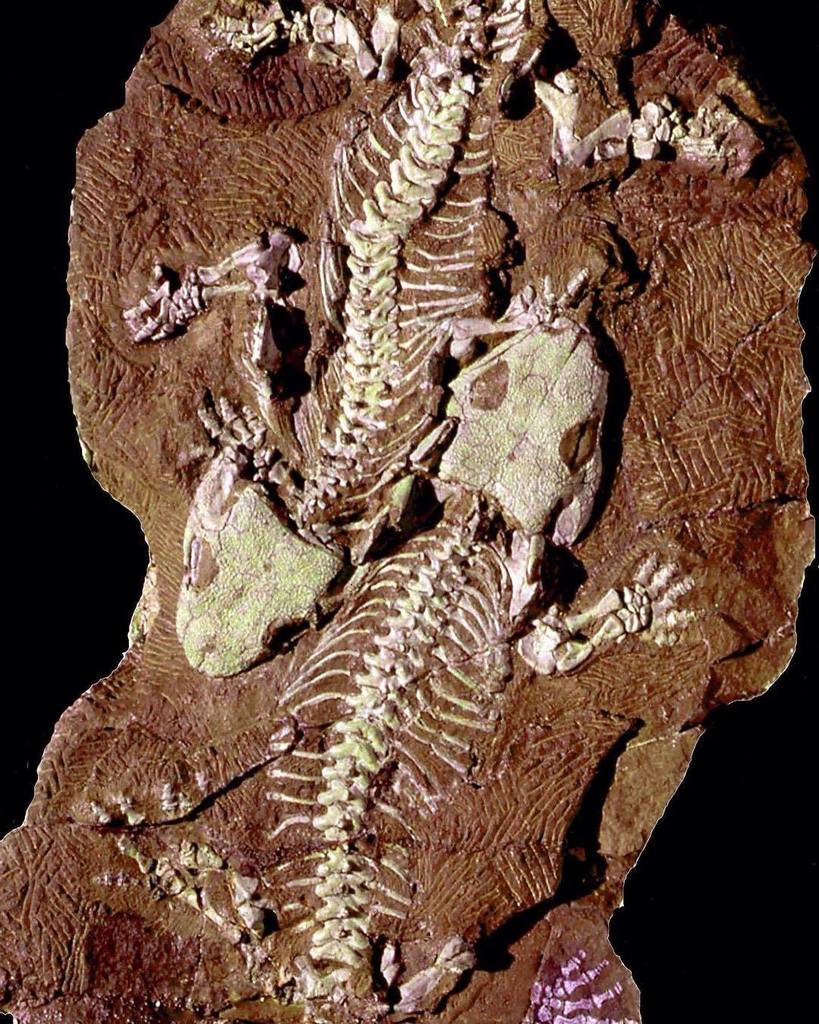
Cacops, is a genus of dissorophid temnospondyls from the Kungurian stage of the early Permian of the United States. Cacops is one of the few olsoniforms whose ontogeny is known. Cacops fossils were almost exclusively known from the Cacops Bone Bed of the Lower Permian Arroyo Formation of Texas for much of the 20th century. New material collected from the Dolese Brothers Quarry, near Richards Spur, Oklahoma in the past few decades has been recovered, painting a clearer picture of what the animal looked and acted like.

Temnospondyli or temnospondyls is a diverse ancient order of small to giant tetrapods—often considered primitive amphibians—that flourished worldwide during the Carboniferous, Permian and Triassic periods, with fossils being found on every continent. A few species continued into the Jurassic and Early Cretaceous periods, but all had gone extinct by the Late Cretaceous. During about 210 million years of evolutionary history, they adapted to a wide range of habitats, including freshwater, terrestrial, and even coastal marine environments. Their life history is well understood, with fossils known from the larval stage, metamorphosis and maturity. Most temnospondyls were semiaquatic, although some were almost fully terrestrial, returning to the water only to breed. These temnospondyls were some of the first vertebrates fully adapted to life on land. Although temnospondyls are amphibians, many had characteristics such as scales and armour-like bony plates that distinguish them from the modern soft-bodied lissamphibians.

Captorhinidae is an extinct family of tetrapods, typically considered primitive reptiles, known from the late Carboniferous to the Late Permian. They had a cosmopolitan distribution across Pangea.

Westlothiana is a genus of reptile-like tetrapod that lived about 338 million years ago during the latest part of the Viséan age of the Carboniferous. Members of the genus bore a superficial resemblance to modern-day lizards. The genus is known from a single species, Westlothiana lizziae. The type specimen was discovered in the East Kirkton Limestone at the East Kirkton Quarry, West Lothian, Scotland in 1984. This specimen was nicknamed "Lizzie the lizard" by fossil hunter Stan Wood, and this name was quickly adopted by other paleontologists and the press. When the specimen was formally named in 1990, it was given the specific name "lizziae" in homage to this nickname. However, despite its similar body shape, Westlothiana is not considered a true lizard. Westlothiana's anatomy contained a mixture of both "labyrinthodont" and reptilian features, and was originally regarded as the oldest known reptile or amniote. However, updated studies have shown that this identification is not entirely accurate. Instead of being one of the first amniotes, Westlothiana was rather a close relative of Amniota. As a result, most paleontologists since the original description place the genus within the group Reptiliomorpha, among other amniote relatives such as diadectomorphs and seymouriamorphs. Later analyses usually place the genus as the earliest diverging member of Lepospondyli, a collection of unusual tetrapods which may be close to amniotes or lissamphibians, or potentially both at the same time.

Seymouriamorpha were a small but widespread group of limbed vertebrates (tetrapods). They have long been considered stem-amniotes (reptiliomorphs), and most paleontologists still accept this point of view, but some analyses suggest that seymouriamorphs are stem-tetrapods. Many seymouriamorphs were terrestrial or semi-aquatic. However, aquatic larvae bearing external gills and grooves from the lateral line system have been found, making them unquestionably amphibians. Though as they matured, they became more terrestrial and reptile-like. They ranged from 30 cm long lizard-sized creatures to the 1.5 m long Enosuchus. If seymouriamorphs are reptiliomorphs, they were the distant relatives of amniotes. Seymouriamorphs form into three main groups, Kotlassiidae, Discosauriscidae, and Seymouriidae, a group that includes the best known genus, Seymouria. The last seymouriamorph became extinct by the end of the Permian.

Platyhystrix is an extinct temnospondyl amphibian with a distinctive sail along its back, similar to the unrelated synapsids, Dimetrodon and Edaphosaurus. It lived during the boundary between the latest Carboniferous and earliest Permian periods throughout what is now known as the Four Corners, Texas, and Kansas about 300 million years ago.

Parareptilia ("near-reptiles") is a subclass or clade of basal sauropsids/reptiles, typically considered the sister taxon to Eureptilia. Parareptiles first arose near the end of the Carboniferous period and achieved their highest diversity during the Permian period. Several ecological innovations were first accomplished by parareptiles among reptiles. These include the first reptiles to return to marine ecosystems (mesosaurs), the first bipedal reptiles, the first reptiles with advanced hearing systems, and the first large herbivorous reptiles. The only parareptiles to survive into the Triassic period were the procolophonoids, a group of small generalists, omnivores, and herbivores. The largest family of procolophonoids, the procolophonids, rediversified in the Triassic, but subsequently declined and became extinct by the end of the period.

Embolomeri is an order of tetrapods or stem-tetrapods, possibly members of Reptiliomorpha. Embolomeres first evolved in the Early Carboniferous (Mississippian) Period and were the largest and most successful predatory tetrapods of the Late Carboniferous (Pennsylvanian) Period. They were specialized semiaquatic predators with long bodies for eel-like undulatory swimming. Embolomeres are characterized by their vertebral centra, which are formed by two cylindrical segments, the pleurocentrum at the rear and intercentrum at the front. These segments are equal in size. Most other tetrapods have pleurocentra and intercentra which are drastically different in size and shape.

Limnoscelis was a genus of large diadectomorph tetrapods from the Late Carboniferous to early Permian of western North America. It includes two species: the type species Limnoscelis paludis from New Mexico, and Limnoscelis dynatis from Colorado, both of which are thought to have lived concurrently. No specimens of Limnoscelis are known from outside of North America. Limnoscelis was carnivorous, and likely semiaquatic, though it may have spent a significant portion of its life on land. Limnoscelis had a combination of derived amphibian and primitive reptilian features, and its placement relative to Amniota has significant implications regarding the origins of the first amniotes.

Kotlassia extinct genus of kotlassiine seymouriamorph from the Late Permian of Russia. The type, and currently only, species is K. prima.

Microleter is an extinct genus of basal procolophonomorph parareptiles which lived in Oklahoma during the Early Permian period. The type and only known species is Microleter mckinzieorum. Microleter is one of several parareptile taxa described from the Richards Spur fissure fills, and can be characterized from its high tooth count, lacrimal/narial contact, short postfrontal, and slit-like temporal emargination edged by the postorbital, jugal, squamosal, and quadratojugal. Contrary to Australothyris, which had a similar phylogenetic position as a basal procolophonomorph, Microleter suggests that early parareptile evolution occurred in Laurasia and that multiple lineages developed openings or emarginations in the temporal region.

The Tambach Formation is an Early Permian-age geologic formation in central Germany. It consists of red to brown-colored sedimentary rocks such as conglomerate, sandstone, and mudstone, and is the oldest portion of the Upper Rotliegend within the Thuringian Forest Basin.

The Organ Rock Formation or Organ Rock Shale is a formation within the late Pennsylvanian to early Permian Cutler Group and is deposited across southeastern Utah, northwestern New Mexico, and northeastern Arizona. This formation notably outcrops around Canyonlands National Park, Natural Bridges National Monument, and Monument Valley of northeast Arizona, southern Utah. The age of the Organ Rock is constrained to the latter half of the Cisuralian epoch by age dates from overlying and underlying formations. Important early terrestrial vertebrate fossils have been recovered from this formation in northern Arizona, southern Utah, and northern New Mexico. These include the iconic Permian terrestrial fauna: Seymouria, Diadectes, Ophiacodon, and Dimetrodon. The fossil assemblage present suggests arid environmental conditions. This is corroborated with paleoclimate data indicative of global drying throughout the early Permian.
This list of fossil amphibians described in 2020 is a list of new taxa of fossil amphibians that were described during the year 2020, as well as other significant discoveries and events related to amphibian paleontology that occurred in 2020.












{kind=link}
{kind=link}
{kind=link}