
Archaeopteryx, sometimes referred to by its German name, "Urvogel", is a genus of avian dinosaurs. The name derives from the ancient Greek ἀρχαῖος (archaīos), meaning "ancient", and πτέρυξ (ptéryx), meaning "feather" or "wing". Between the late 19th century and the early 21st century, Archaeopteryx was generally accepted by palaeontologists and popular reference books as the oldest known bird. Older potential avialans have since been identified, including Anchiornis, Xiaotingia, and Aurornis.

The chukar partridge, or simply chukar, is a Palearctic upland gamebird in the pheasant family Phasianidae. It has been considered to form a superspecies complex along with the rock partridge, Philby's partridge and Przevalski's partridge and treated in the past as conspecific particularly with the first. This partridge has well-marked black and white bars on the flanks and a black band running from the forehead across the eye down the head to form a necklace that encloses a white throat. Native to Asia, the species has been introduced into many other places and feral populations have established themselves in parts of North America and New Zealand. This bird can be found in parts of Middle East and temperate Asia.

Microraptor is a genus of small, four-winged dromaeosaurid dinosaurs. Numerous well-preserved fossil specimens have been recovered from Liaoning, China. They date from the early Cretaceous Jiufotang Formation, 125 to 120 million years ago. Three species have been named, though further study has suggested that all of them represent variation in a single species, which is properly called M. zhaoianus. Cryptovolans, initially described as another four-winged dinosaur, is usually considered to be a synonym of Microraptor.

Maniraptora is a clade of coelurosaurian dinosaurs which includes the birds and the non-avian dinosaurs that were more closely related to them than to Ornithomimus velox. It contains the major subgroups Avialae, Dromaeosauridae, Troodontidae, Oviraptorosauria, and Therizinosauria. Ornitholestes and the Alvarezsauroidea are also often included. Together with the next closest sister group, the Ornithomimosauria, Maniraptora comprises the more inclusive clade Maniraptoriformes. Maniraptorans first appear in the fossil record during the Jurassic Period, and survive today as living birds.

Bird flight is the primary mode of locomotion used by most bird species in which birds take off and fly. Flight assists birds with feeding, breeding, avoiding predators, and migrating.

Scansoriopterygidae is an extinct family of climbing and gliding maniraptoran dinosaurs. Scansoriopterygids are known from five well-preserved fossils, representing four species, unearthed in the Tiaojishan Formation fossil beds of Liaoning and Hebei, China.
The evolution of birds began in the Jurassic Period, with the earliest birds derived from a clade of theropod dinosaurs named Paraves. Birds are categorized as a biological class, Aves. For more than a century, the small theropod dinosaur Archaeopteryx lithographica from the Late Jurassic period was considered to have been the earliest bird. Modern phylogenies place birds in the dinosaur clade Theropoda. According to the current consensus, Aves and a sister group, the order Crocodilia, together are the sole living members of an unranked reptile clade, the Archosauria. Four distinct lineages of bird survived the Cretaceous–Paleogene extinction event 66 million years ago, giving rise to ostriches and relatives (Palaeognathae), ducks and relatives (Anseriformes), ground-living fowl (Galliformes), and "modern birds" (Neoaves).

Crested arguses are large and spectacular peafowl-like birds in the genus Rheinardia of the pheasant family.
The scientific question of within which larger group of animals birds evolved has traditionally been called the "origin of birds". The present scientific consensus is that birds are a group of maniraptoran theropod dinosaurs that originated during the Mesozoic Era.
Around 350 BCE, Aristotle and other philosophers of the time attempted to explain the aerodynamics of avian flight. Even after the discovery of the ancestral bird Archaeopteryx which lived over 150 million years ago, debates still persist regarding the evolution of flight. There are three leading hypotheses pertaining to avian flight: Pouncing Proavis model, Cursorial model, and Arboreal model.

Paraves are a widespread group of theropod dinosaurs that originated in the Middle Jurassic period. In addition to the extinct dromaeosaurids, troodontids, anchiornithids, and possibly the scansoriopterygids, the group also contains the avialans, which include diverse extinct taxa as well as the over 10,000 species of living birds. Basal members of Paraves are well known for the possession of an enlarged claw on the second digit of the foot, which was held off the ground when walking in some species. A number of differing scientific interpretations of the relationships between paravian taxa exist. New fossil discoveries and analyses make the classification of Paraves an active subject of research.
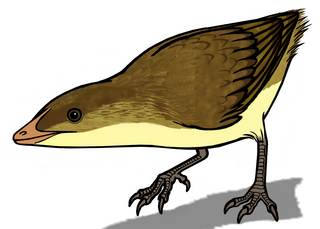
Eoalulavis is a monotypic genus of enantiornithean bird that lived during the Barremian, in the Lower Cretaceous around 125 million years ago. The only known species is Eoalulavis hoyasi.

Anchiornis is a genus of small, four-winged paravian dinosaurs, with only one known species, the type species Anchiornis huxleyi, named for its similarity to modern birds. The Latin name Anchiornis derives from a Greek word meaning "near bird", and huxleyi refers to Thomas Henry Huxley, a contemporary of Charles Darwin.
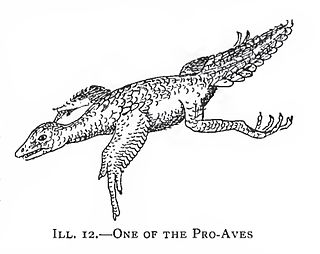
Proavis refers to a hypothetical extinct species or hypothetical extinct taxon and was coined in the early 20th century in an attempt to support and explain the hypothetical evolutionary steps and anatomical adaptations leading from non-avian theropod dinosaurs to birds. The term has also been used by defenders of the thecodontian origin of birds.

Certain species of fish and birds are able to locomote in both air and water, two fluid media with very different properties. A fluid is a particular phase of matter that deforms under shear stresses and includes any type of liquid or gas. Because fluids are easily deformable and move in response to applied forces, efficiently locomoting in a fluid medium presents unique challenges. Specific morphological characteristics are therefore required in animal species that primarily depend on fluidic locomotion. Because the properties of air and water are so different, swimming and flying have very disparate morphological requirements. As a result, despite the large diversity of animals that are capable of flight or swimming, only a limited number of these species have mastered the ability to both fly and swim. These species demonstrate distinct morphological and behavioral tradeoffs associated with transitioning from air to water and water to air.

The Origin of Birds is an early synopsis of bird evolution written in 1926 by Gerhard Heilmann, a Danish artist and amateur zoologist. The book was born from a series of articles published between 1913 and 1916 in Danish, and although republished as a book it received mainly criticism from established scientists and got little attention within Denmark. The English edition of 1926, however, became highly influential at the time due to the breadth of evidence synthesized as well as the artwork used to support its arguments. It was considered the last word on the subject of bird evolution for several decades after its publication.

Yi is a genus of scansoriopterygid dinosaurs from the Late Jurassic of China. Its only species, Yi qi, is known from a single fossil specimen of an adult individual found in Middle or Late Jurassic Tiaojishan Formation of Hebei, China, approximately 159 million years ago. It was a small, possibly tree-dwelling (arboreal) animal. Like other scansoriopterygids, Yi possessed an unusual, elongated third finger, that appears to have helped to support a membranous gliding plane made of skin. The planes of Yi qi were also supported by a long, bony strut attached to the wrist. This modified wrist bone and membrane-based plane is unique among all known dinosaurs, and might have resulted in wings similar in appearance to those of bats.

Zhenyuanlong is a genus of dromaeosaurid dinosaur from the Yixian Formation of Liaoning, China. It lived during the Aptian age of the early Cretaceous period, approximately 125 million years ago. It is known from a single specimen belonging to the species Zhenyuanlong suni. This type specimen preserved a nearly complete skeleton that contains traces of feathers, including long tail feathers and large wings. In addition to further complicating diversity of Liaoning dromaeosaurids, this specimen provides the first evidence of well-developed pennaceous feathers in a large, non-flying dromaeosaur, raising the question of what function such wings would serve.

Feathers: The Evolution of a Natural Miracle is a natural history book by American conservation biologist Thor Hanson. Published by Basic Books in 2011 and written for general audiences, the book discusses the significance of feathers, their evolution, and their history both in nature and in use by humans.
Feather development occurs in the epidermal layer of the skin in birds. It is a complicated process involving many steps. Once the feathers are fully developed, there are six different types of feathers: contour, flight, down, filoplumes, semiplumes, and bristle feathers. Feathers were not originally meant for flight. The exact reason why feathers evolved is still unknown. Birds are thought to be descendants of dinosaurs and new technology using melanosomes found in dinosaur fossils has shown that certain dinosaurs that could not fly had feathers.
