A blood type is a classification of blood, based on the presence and absence of antibodies and inherited antigenic substances on the surface of red blood cells (RBCs). These antigens may be proteins, carbohydrates, glycoproteins, or glycolipids, depending on the blood group system. Some of these antigens are also present on the surface of other types of cells of various tissues. Several of these red blood cell surface antigens can stem from one allele and collectively form a blood group system.
Rh disease is a type of hemolytic disease of the fetus and newborn (HDFN). HDFN due to anti-D antibodies is the proper and currently used name for this disease as the Rh blood group system actually has more than 50 antigens and not only the D-antigen. The term "Rh Disease" is commonly used to refer to HDFN due to anti-D antibodies, and prior to the discovery of anti-Rho(D) immune globulin, it was the most common type of HDFN. The disease ranges from mild to severe, and occurs in the second or subsequent pregnancies of Rh-D negative women when the biologic father is Rh-D positive.

Duffy antigen/chemokine receptor (DARC), also known as Fy glycoprotein (FY) or CD234, is a protein that in humans is encoded by the ACKR1 gene.

α-Galactosidase is a glycoside hydrolase enzyme that catalyses the following reaction:
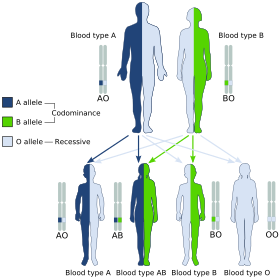
The term human blood group systems is defined by the International Society of Blood Transfusion (ISBT) as systems in the human species where cell-surface antigens—in particular, those on blood cells—are "controlled at a single gene locus or by two or more very closely linked homologous genes with little or no observable recombination between them", and include the common ABO and Rh (Rhesus) antigen systems, as well as many others; 44 human systems are identified as of December 2022.
The Kell antigen system is a human blood group system, that is, a group of antigens on the human red blood cell surface which are important determinants of blood type and are targets for autoimmune or alloimmune diseases which destroy red blood cells. The Kell antigens are K, k, Kpa, Kpb, Jsa and Jsb. The Kell antigens are peptides found within the Kell protein, a 93-kilodalton transmembrane zinc-dependent endopeptidase which is responsible for cleaving endothelin-3.

The Rh blood group system is a human blood group system. It contains proteins on the surface of red blood cells. After the ABO blood group system, it is the most likely to be involved in transfusion reactions. The Rh blood group system consisted of 49 defined blood group antigens in 2005. As of 2023, there are over 50 antigens among which the five antigens D, C, c, E, and e are the most important. There is no d antigen. Rh(D) status of an individual is normally described with a positive (+) or negative (−) suffix after the ABO type. The terms Rh factor, Rh positive, and Rh negative refer to the Rh(D) antigen only. Antibodies to Rh antigens can be involved in hemolytic transfusion reactions and antibodies to the Rh(D) and Rh antigens confer significant risk of hemolytic disease of the newborn.
Animal erythrocytes have cell surface antigens that undergo polymorphism and give rise to blood types. Antigens from the human ABO blood group system are also found in apes and Old World monkeys, and the types trace back to the origin of humanoids. Other animal blood sometimes agglutinates with human blood group reagents, but the structure of the blood group antigens in animals is not always identical to those typically found in humans. The classification of most animal blood groups therefore uses different blood typing systems to those used for classification of human blood.
The MNS antigen system is a human blood group system based upon two genes on chromosome 4. There are currently 50 antigens in the system, but the five most important are called M, N, S, s, and U.
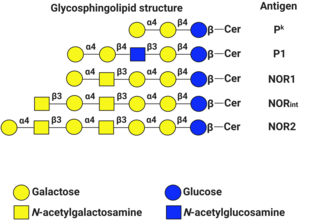
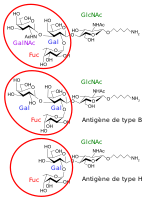
P1PK is a human blood group system based upon the A4GALT gene on chromosome 22. The P antigen was first described by Karl Landsteiner and Philip Levine in 1927. The P1PK blood group system consists of three glycosphingolipid antigens: Pk, P1 and NOR. In addition to glycosphingolipids, terminal Galα1→4Galβ structures are present on complex-type N-glycans. The GLOB antigen is now the member of the separate GLOB (globoside) blood group system.
The Lewis antigen system is a human blood group system. It is based upon two genes on chromosome 19: FUT3, or Lewis gene; and FUT2, or Secretor gene. Both genes are expressed in glandular epithelia. FUT2 has a dominant allele which codes for an enzyme and a recessive allele which does not produce a functional enzyme. Similarly, FUT3 has a functional dominant allele (Le) and a non-functional recessive allele (le).

The Ii antigen system is a human blood group system based upon a gene on chromosome 6 and consisting of the I antigen and the i antigen. The I antigen is normally present on the cell membrane of red blood cells in all adults, while the i antigen is present in fetuses and newborns.

The Diego antigen system is composed of 21 blood factors or antigens carried on the Band 3 glycoprotein, also known as Anion Exchanger 1 (AE1). The antigens are inherited through various alleles of the gene SLC4A1, located on human chromosome 17. The AE1 glycoprotein is expressed only in red blood cells and, in a shortened form, in some cells in the kidney. The Diegoa antigen is fairly common in Indigenous peoples of the Americas and East Asians, but very rare or absent in most other populations, supporting the theory that the two groups share common ancestry.

Glycophorin A (MNS blood group), also known as GYPA, is a protein which in humans is encoded by the GYPA gene. GYPA has also recently been designated CD235a (cluster of differentiation 235a).
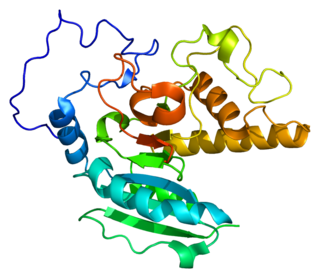
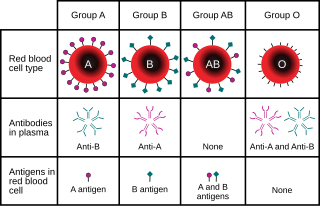
Histo-blood group ABO system transferase is an enzyme with glycosyltransferase activity, which is encoded by the ABO gene in humans. It is ubiquitously expressed in many tissues and cell types. ABO determines the ABO blood group of an individual by modifying the oligosaccharides on cell surface glycoproteins. Variations in the sequence of the protein between individuals determine the type of modification and the blood group. The ABO gene also contains one of 27 SNPs associated with increased risk of coronary artery disease.
Isoantibodies, formerly called alloantibodies, are antibodies produced by an individual against isoantigens produced by members of the same species. In the case of the species Homo sapiens, for example, there are a significant number of antigens that are different in every individual. When antigens from another individual are introduced into another's body, these isoantibodies immediately bind to and destroy them.
Cis AB is a type of rare mutation in the ABO gene. It happens when the transferase allele contains a mix of amino acids from either A or B alleles, producing a bifunctional enzyme that can produce both types of antigens, usually with one weaker than the other. This results in a serum test result much like the standard, separate (trans) AB phenotype, although the weaker antigen can occasionally fail to be detected. It complicates the basic inheritance pattern and blood-transfusion compatibility matching for ABO blood typing.

Blood compatibility testing is conducted in a medical laboratory to identify potential incompatibilities between blood group systems in blood transfusion. It is also used to diagnose and prevent some complications of pregnancy that can occur when the baby has a different blood group from the mother. Blood compatibility testing includes blood typing, which detects the antigens on red blood cells that determine a person's blood type; testing for unexpected antibodies against blood group antigens ; and, in the case of blood transfusions, mixing the recipient's plasma with the donor's red blood cells to detect incompatibilities (crossmatching). Routine blood typing involves determining the ABO and RhD type, and involves both identification of ABO antigens on red blood cells and identification of ABO antibodies in the plasma. Other blood group antigens may be tested for in specific clinical situations.
The Lan blood group system is a human blood group defined by the presence or absence of the Lan antigen on a person's red blood cells. More than 99.9% of people are positive for the Lan antigen. Individuals with the rare Lan-negative blood type, which is a recessive trait, can produce an anti-Lan antibody when exposed to Lan-positive blood. Anti-Lan antibodies may cause transfusion reactions on subsequent exposures to Lan-positive blood, and have also been implicated in mild cases of hemolytic disease of the newborn. However, the clinical significance of the antibody is variable. The antigen was first described in 1961, and Lan was officially designated a blood group in 2012.
The Sid blood group system is a human blood group defined by the presence or absence of the Sd(a) antigen on a person's red blood cells. About 96% of people are positive for the Sd(a) antigen, which is inherited as a dominant trait. Among Sd(a) positive individuals, the expression of the antigen ranges from extremely weak to extremely strong. Very strong expression of the antigen is referred to as a Sd(a++) phenotype. In addition to being expressed on red blood cells, Sd(a) is secreted in bodily fluids such as saliva and breast milk, and is found in the highest concentrations in urine. Urine testing is considered the most reliable method for determining a person's Sid blood type.