The Rh blood group system is a human blood group system. It contains proteins on the surface of red blood cells. After the ABO blood group system, it is the most likely to be involved in transfusion reactions. The Rh blood group system consisted of 49 defined blood group antigens[1] in 2005. As of 2023,[update] there are over 50 antigens among which the five antigens D, C, c, E, and e are the most important. There is no d antigen. Rh(D) status of an individual is normally described with a positive (+) or negative (−) suffix after the ABO type (e.g., someone who is A+ has the A antigen and Rh(D) antigen, whereas someone who is A− has the A antigen but lacks the Rh(D) antigen). The terms Rh factor, Rh positive, and Rh negative refer to the Rh(D) antigen only. Antibodies to Rh antigens can be involved in hemolytic transfusion reactions and antibodies to the Rh(D) and Rh antigens confer significant risk of hemolytic disease of the newborn.
The Rh blood group system has two sets of nomenclatures: one developed by Ronald Fisher and R. R. Race, the other by Wiener. Both systems reflected alternative theories of inheritance. The Fisher–Race system, which is more commonly in use today, uses the CDE nomenclature. This system was based on the theory that a separate gene controls the product of each corresponding antigen (e.g., a "D gene" produces D antigen, and so on). However, the d gene was hypothetical, not actual.
The Wiener system used the Rh–Hr nomenclature. This system was based on the theory that there was one gene at a single locus on each of the two copies of chromosome 1, each contributing to production of multiple antigens. In this theory, a gene R1 is supposed to give rise to the "blood factors" Rh0, rh′, and rh″ (corresponding to modern nomenclature of the D, C, and E antigens) and the gene r to produce hr′ and hr″ (corresponding to modern nomenclature of the c and e antigens).[3]
Notations of the two theories are used interchangeably in blood banking (e.g., Rho(D) meaning RhD positive). Wiener's notation is more complex and cumbersome for routine use. Because it is simpler to explain, the Fisher–Race theory has become more widely used.[citation needed]
DNA testing has shown that both are partially correct: There are in fact two linked genes, the RHD gene which produces a single immune specificity (anti-D) and the RHCE gene with multiple specificities (anti-C, anti-c, anti-E, anti-e). Thus, Wiener's postulate that a gene could have multiple specificities (something many did not give credence to originally) has been proved to be correct. On the other hand, Wiener's theory that there is only one gene has proved to be incorrect, as has the Fisher–Race theory that there are three genes, rather than the two. The CDE notation used in the Fisher–Race nomenclature is sometimes rearranged to DCE to more accurately represent the co-location of the C and E encoding on the RhCE gene, and to make interpretation easier.[citation needed]
Antigens
The proteins which carry the Rh antigens are transmembrane proteins, whose structure suggests that they are ion channels.[4] The main antigens are D, C, E, c and e, which are encoded by two adjacent gene loci, the RHD gene which encodes the RhD protein with the D antigen (and variants)[5] and the RHCE gene which encodes the RhCE protein with the C, E, c and e antigens (and variants).[6] There is no d antigen. Lowercase "d" indicates the absence of the D antigen (the gene is usually deleted or otherwise nonfunctional).[citation needed]
1. This is the Rh-positive blood cell. 2. This is the Rh-negative blood cell. 3. These are the antigens on the Rh-positive blood cell that make it positive. The antigens allow the positive blood cell to attach to specific antibodies.
Rh phenotypes are readily identified through the presence or absence of the Rh surface antigens. As can be seen in the table below, most of the Rh phenotypes can be produced by several different Rh genotypes. The exact genotype of any individual can only be identified by DNA analysis. Regarding patient treatment, only the phenotype is usually of any clinical significance to ensure a patient is not exposed to an antigen they are likely to develop antibodies against. A probable genotype may be speculated on, based upon the statistical distributions of genotypes in the patient's place of origin.[citation needed]
R0 (cDe or Dce) is today most common in Africa. The allele was thus often assumed in early blood group analyses to have been typical of populations on the continent; particularly in areas below the Sahara. Ottensooser et al. (1963) suggested that high R0 frequencies were likely characteristic of the ancient Judean Jews, who had emigrated from Egypt prior to their dispersal throughout the Mediterranean Basin and Europe[7] on the basis of high R0 percentages among Sephardi and AshkenaziJews compared to native European populations and the relative genetic isolation of Ashkenazim. However, more recent studies have found R0 frequencies as low as 24.3% among some Afroasiatic-speaking groups in the Horn of Africa,[8] as well as higher R0 frequencies among certain other Afroasiatic speakers in North Africa (37.3%)[9] and among some Palestinians in the Levant (30.4%).[10] On the contrary, at a frequency of 47.2% of the population of Basque country having the lack of the D antigen, these people display the highest frequency of the Rh negative phenotype.[11]
Rh phenotypes and genotypes (UK, 1948)
Phenotype expressed on cell
Genotype expressed in DNA
Prevalence (%)
Fisher–Race notation
Wiener notation
D+ C+ E+ c+ e+ (RhD+)
Dce/DCE
R0RZ
0.0125
Dce/dCE
R0rY
0.0003
DCe/DcE
R1R2
11.8648
DCe/dcE
R1r″
0.9992
DcE/dCe
R2r′
0.2775
DCE/dce
RZr
0.1893
D+ C+ E+ c+ e− (RhD+)
DcE/DCE
R2RZ
0.0687
DcE/dCE
R2rY
0.0014
DCE/dcE
RZr″
0.0058
D+ C+ E+ c− e+ (RhD+)
DCe/dCE
R1rY
0.0042
DCE/dCe
RZr′
0.0048
DCe/DCE
R1RZ
0.2048
D+ C+ E+ c− e− (RhD+)
DCE/DCE
RZRZ
0.0006
DCE/dCE
RZrY
< 0.0001
D+ C+ E− c+ e+ (RhD+)
Dce/dCe
R0r′
0.0505
DCe/dce
R1r
32.6808
DCe/Dce
R1R0
2.1586
D+ C+ E− c− e+ (RhD+)
DCe/DCe
R1R1
17.6803
DCe/dCe
R1r′
0.8270
D+ C− E+ c+ e+ (RhD+)
DcE/Dce
R2R0
0.7243
Dce/dcE
R0r″
0.0610
DcE/dce
R2r
10.9657
D+ C− E+ c+ e− (RhD+)
DcE/DcE
R2R2
1.9906
DcE/dcE
R2r″
0.3353
D+ C− E− c+ e+ (RhD+)
Dce/Dce
R0R0
0.0659
Dce/dce
R0r
1.9950
D− C+ E+ c+ e+ (RhD−)
dce/dCE
rrY
0.0039
dCe/dcE
r′r″
0.0234
D− C+ E+ c+ e− (RhD−)
dcE/dCE
r″rY
0.0001
D− C+ E+ c− e+ (RhD−)
dCe/dCE
r′rY
0.0001
D− C+ E+ c− e− (RhD−)
dCE/dCE
rYrY
< 0.0001
D− C+ E− c+ e+ (RhD−)
dce/dCe
rr′
0.7644
D− C+ E− c− e+ (RhD−)
dCe/dCe
r′r′
0.0097
D− C− E+ c+ e+ (RhD−)
dce/dcE
rr″
0.9235
D− C− E+ c+ e− (RhD−)
dcE/dcE
r″r″
0.0141
D− C− E− c+ e+ (RhD−)
dce/dce
rr
15.1020
• Figures taken from a study performed in 1948 on a sample of 2000 people in the United Kingdom.[12]
Rh phenotypes in patients and donors in Turkey[13]
Rh Phenotype
CDE
Patients (%)
Donors (%)
R1r
CcDe
37.4
33.0
R1R2
CcDEe
35.7
30.5
R1R1
CDe
5.7
21.8
rr
ce
10.3
11.6
R2r
cDEe
6.6
10.4
R0R0
cDe
2.8
2.7
R2R2
cDE
2.8
2.4
rr″
cEe
–
0.98
RZRZ
CDE
–
0.03
rr′
Cce
0.8
–
Rh antibodies
Rh antibodies are Immunoglobulin G (IgG) antibodies which are acquired through exposure to Rh-positive blood (generally either through pregnancy or transfusion of blood products). The D antigen is the most immunogenic of all the non-ABO antigens. Approximately 80% of individuals who are D-negative and exposed to a single D-positive unit will produce an anti-D antibody. The percentage of alloimmunization is significantly reduced in patients who are actively exsanguinating.[14]
All Rh antibodies except D display dosage (antibody reacts more strongly with red cells homozygous for an antigen than cells heterozygous for the antigen (EE stronger reaction vs Ee)).
If anti-E is detected, the presence of anti-c should be strongly suspected (due to combined genetic inheritance). It is therefore common to select c-negative and E-negative blood for transfusion patients who have an anti-E and lack the c antigen (in general, a patient will not produce antibodies against their own antigens). Anti-c is a common cause of delayed hemolytic transfusion reactions.[15]
The hemolytic condition occurs when there is an incompatibility between the blood types of the mother and fetus. There is also potential incompatibility if the mother is Rh negative and the father is positive. When the mother conceives for the first time, with a positive child, she will become extremely sensitive. When any incompatibility is detected when she conceives the second time in less than two years then, the mother often receives an injection at 28 weeks gestation and at birth to avoid the development of antibodies towards the fetus. If not given, then the baby will be dead and must be aborted. These terms do not indicate which specific antigen-antibody incompatibility is implicated. The disorder in the fetus due to Rh D incompatibility is known as erythroblastosis fetalis.
Hemolytic comes from two words: "hema" (blood) and "lysis" (solution) or breaking down of red blood cells
Erythroblastosis refers to the making of immature red blood cells
Fetalis refers to the fetus.
When the condition is caused by the Rh D antigen-antibody incompatibility, it is called Rh D Hemolytic disease of the newborn or Rh disease. Here, sensitization to Rh D antigens (usually by feto-maternal transfusion during pregnancy) may lead to the production of maternal IgG anti-D antibodies which can pass through the placenta. This is of particular importance to D negative females at or below childbearing age, because any subsequent pregnancy may be affected by the Rh D hemolytic disease of the newborn if the baby is D positive. The vast majority of Rh disease is preventable in modern antenatal care by injections of IgG anti-D antibodies (Rho(D) Immune Globulin). The incidence of Rh disease is mathematically related to the frequency of D negative individuals in a population, so Rh disease is rare in old-stock populations of Africa and the eastern half of Asia, and the Indigenous peoples of Oceania and the Americas, but more common in other genetic groups, most especially Western Europeans, but also other West Eurasians, and to a lesser degree, native Siberians, as well as those of mixed-race with a significant or dominant descent from those (e.g. the vast majority of Latin Americans and Central Asians).
Symptoms and signs in the fetus:
Enlarged liver, spleen, or heart and fluid buildup in the fetus' abdomen seen via ultrasound.
Symptoms and signs in the newborn:
Anemia that creates the newborn's pallor (pale appearance).
Jaundice or yellow discoloration of the newborn's skin, sclera or mucous membrane. This may be evident right after birth or after 24–48 hours after birth. This is caused by bilirubin (one of the end products of red blood cell destruction).
Enlargement of the newborn's liver and spleen.
The newborn may have severe edema of the entire body.
Dyspnea (difficulty breathing)
Other animals with Rh-like antigens causing hemolytic disease of the newborn
Rh disease only occurs in human fetuses however a similar disease called Neonatal isoerythrolysis (NI) can be observed in animal species of newborn horses, mules, pigs, cats, cattle, and dogs. What differs between Rh disease and NI is the pathogenesis of hemolysis between human fetuses and the animal species. With human mothers, the maternal antibodies are formed from the sensitization of foreign antigens of her unborn fetus’s red blood cells passing through the placenta causing hemolysis before birth, with other animals however, these maternal antibodies are not passed through the placenta but through colostrum. The newborn animal is without NI but soon develops hemolytic anemia after initial ingestion of its mother’s colostrum that contain antibodies that can be absorbed through the newborn’s intestines and are incompatible to its red blood cell antigen. After 48 hours of birth, the newborn may be allowed to nurse from its mother as her antibodies can no longer be absorbed through the neonate’s intestines. Because the most active newborn animals consume the most colostrum, they may be the ones who are most affected by the blood incompatibility of antigen and antibody.[16]
According to a comprehensive study, the worldwide frequency of Rh-positive and Rh-negative blood types is approximately 94% and 6%, respectively. The same study concluded that the share of the population with Rh-negative blood type is set to fall further in the future primarily due to low population growth in Europe.[17] The frequency of Rh factor blood types and the RhD neg allele gene differs in various populations.[citation needed]
Population data for the Rh D factor and RhD neg allele[18]
This is a Punnett square for Rh factor inheritance. This square specifically shows two heterozygous Rh positive parents and the possible genotypes/phenotypes the offspring could have.
The D antigen is inherited as one gene (RHD) (on the short arm of the first chromosome, p36.13–p34.3) with various alleles. Typically, Rhesus positive people have an intact RHD gene while negative people lack the gene (or have mutations in it). However, there are exceptions: for instance, Japanese and black Africans may have an intact gene that is not expressed or only at very low levels.[27] The gene codes for the RhD protein on the red blood cell membrane. D− individuals who lack a functional RHD gene do not produce the D antigen, and may be immunized by D+ blood.[citation needed]
The D antigen is a dominant trait. If both of a child's parents are Rh negative, the child will definitely be Rh negative. Otherwise the child may be Rh positive or Rh negative, depending on the parents' specific genotypes.[28]
The epitopes for the next 4 most common Rh antigens, C, c, E and e are expressed on the highly similar RhCE protein that is genetically encoded in the RHCE gene, also found on chromosome 1. It has been shown that the RHD gene arose by duplication of the RHCE gene during primate evolution. Mice have just one RH gene.[29]
The RHAG gene, which is responsible for encoding Rh-associated glycoprotein (RhAG), is found on chromosome 6a.
The polypeptides produced from the RHD and RHCE genes form a complex on the red blood cell membrane with the Rh-associated glycoprotein.[15]
On the basis of structural homology it has been proposed that the product of RHD gene, the RhD protein, is a membrane transport protein of uncertain specificity (CO2 or NH3) and unknown physiological role.[30][31] The three-dimensional structure of the related RHCG protein and biochemical analysis of the RhD protein complex indicates that the RhD protein is one of three subunits of an ammonia transporter.[32][33] Three recent studies[34][35][36] have reported a protective effect of the RhD-positive phenotype, especially RhD heterozygosity, against the negative effect of latent toxoplasmosis on psychomotor performance in infected subjects. RhD-negative compared to RhD-positive subjects without anamnestic titres of anti-Toxoplasma antibodies have shorter reaction times in tests of simple reaction times. And conversely, RhD-negative subjects with anamnestic titres (i.e. with latent toxoplasmosis) exhibited much longer reaction times than their RhD-positive counterparts. The published data suggested that only the protection of RhD-positive heterozygotes was long term in nature; the protection of RhD-positive homozygotes decreased with duration of the infection while the performance of RhD-negative homozygotes decreased immediately after the infection. The overall change in reaction times was always larger in the RhD-negative group than in the RhD-positive.[citation needed]
Non-human Rh proteins
This section may require cleanup to meet Wikipedia's quality standards. The specific problem is: Should not be here, where we deal with the non-glycosylated variable Rh antigens; there should be an article describing all the Rh proteins (including RHAG, RHBG, RHCG, RHD, RHCE]], and that's where it should go. Please help improve this section if you can.(January 2024) ( Learn how and when to remove this message )
Rh-like proteins can be found even in species other than vertebrates (which have red blood cells) – worms, bacteria, and algae. All these Rh proteins have the same biochemical function of transporting CO2, differing slightly in their amino acid sequences. The Rh family as a whole is related to ammonia transporters (Amt). In C. elegans worms, disruption of the Rh1 gene causes growth defects under high CO2 levels. Chlamydomonas reinhardtii algae fail to grow rapidly if its Rh gene is knocked down. Although in vitro evidence shows the Rh complex is capable of moving ammonia, its disruption does not cause growth defects under modified ammonia levels.[37][38]
RHD polymorphism
Origin of RHD polymorphism
For a long time, the origin of RHD polymorphism was an evolutionary enigma.[39][40][41] Before the advent of modern medicine, the carriers of the rarer allele (e.g. RhD-negative women in a population of RhD positives or RhD-positive men in a population of RhD negatives) were at a disadvantage as some of their children (RhD-positive children born to preimmunised RhD-negative mothers) were at a higher risk of fetal or newborn death or health impairment from hemolytic disease.[citation needed]
Natural selection aside, the RHD-RHCE region is structurally predisposed to many mutations seen in humans, since the pair arose by gene duplication and remain similar enough for unequal crossing over to occur.[29] In addition to the case where D is deleted, crossover can also produce a single gene mixing exons from both RHD and RHCE, forming the majority of partial D types.[42]:323
In serologic testing, D positive blood is easily identified. Units that are D negative are often retested to rule out a weaker reaction. This was previously referred to as Du, which has been replaced.[42]:322 By definition, weak D phenotype is characterized by negative reaction with anti-D reagent at immediate spin (IS), negative reaction after 37°C incubation, and positive reaction at anti-human globulin (AHG) phase. Weak D phenotype can occur in several ways. In some cases, this phenotype occurs because of an altered surface protein that is more common in people of European descent. An inheritable form also occurs, as a result of a weakened form of the R0 gene. Weak D may also occur as "C in trans", whereby a C gene is present on the opposite chromosome to a D gene (as in the combination R0r', or "Dce/dCe"). The testing is difficult, since using different anti-D reagents, especially the older polyclonal reagents, may give different results.
The practical implication of this is that people with this sub-phenotype will have a product labeled as "D positive" when donating blood. When receiving blood, they are sometimes typed as a "D negative", though this is the subject of some debate. Most "Weak D" patients can receive "D positive" blood without complications.[42]:323 However, it is important to correctly identify the ones that have to be considered D+ or D−. This is important, since most blood banks have a limited supply of "D negative" blood and the correct transfusion is clinically relevant. In this respect, genotyping of blood groups has much simplified this detection of the various variants in the Rh blood group system.
Partial D
It is important to differentiate weak D (due to a quantitative difference in the D antigen) from partial D (due to a qualitative difference in the D antigen). Simply put, the weak D phenotype is due to a reduced number of D antigens on a red blood cell. In contrast, the partial D phenotype is due to an alteration in D-epitopes. Thus, in partial D, the number of D antigens is not reduced but the protein structure is altered. These individuals, if alloimmunized to D, can produce an anti-D antibody. Therefore, partial D patients who are donating blood should be labeled as D-positive but, if receiving blood, they should be labeled as D-negative and receive D-negative units.[15]
In the past, partial D was called 'D mosaic' or 'D variant.' Different partial D phenotypes are defined by different D epitopes on the outer surface of the red blood cell membrane. More than 30 different partial D phenotypes have been described.[15]
Rhnull phenotype
Rhnull individuals have no Rh antigens (no Rh or RhAG) on their red blood cells.[43] This rare condition[43] has been called "Golden Blood".[44] As a consequence of Rh antigen absence, Rhnull red blood cells also lack LW and Fy5 and show weak expression of S, s, and U antigens.
Red blood cells lacking Rh/RhAG proteins have structural abnormalities (such as stomatocytosis) and cell membrane defects that can result in hemolytic anemia.[15][43]
The first Rhnull blood was discovered in an Aboriginal Australian woman, in 1961.[45] Only 43 individuals have been reported to have it worldwide. Only nine active donors have been reported.[44] Its properties make it attractive in numerous medical applications, but scarcity makes it expensive to transport and acquire.[46]
Other Rh group antigens
As of 2023, over 50 antigens have been described in the Rh group system; among those described here, the D, C, c, E and e antigens are the most important. The others are much less frequently encountered or are rarely clinically significant. Each is given a number, though the highest assigned number (CEVF or RH61 according to the ISBT terminology) is not an accurate reflection of the antigens encountered since many (e.g. Rh38) have been combined, reassigned to other groups, or otherwise removed.[42]:324
Some of the other Rh "antigens" are f ("ce", RH6), Ce (RH7), Cw (RH8), Cx (RH9), V (RH10), Ew (RH11), G (RH12), Tar (RH40), VS (RH20), Dw (RH23), and CE (RH22). Some of these groups, including f, Ce and CE, describe grouping of some existing groups. Others, like V, describe an epitope created by some other mutation on the RHD and RHCE genes. V in particular is caused by a mutation on RHCE.[47]
History
The term "Rh" was originally an abbreviation of "Rhesus factor". It was discovered in 1939 by Karl Landsteiner and Alexander S. Wiener, who, at the time, believed it to be a similar antigen found in rhesus macaque red blood cells. It was subsequently discovered that the human factor is not identical to the rhesus monkey factor, but by then, "Rhesus Group" and like terms were already in widespread, worldwide use. Thus, notwithstanding it is a misnomer, the term survives (e.g., rhesus blood group system and the obsolete terms rhesus factor, rhesus positive, and rhesus negative – all three of which actually refer specifically and only to the Rh D factor and are thus misleading when unmodified). Contemporary practice is to use "Rh" as a term of art instead of "Rhesus" (e.g., "Rh Group", "Rh factors", "Rh D", etc.).
The significance of their discovery was not immediately apparent and was only realized in 1940, after subsequent findings by Philip Levine and Rufus Stetson.[48] The serum that led to the discovery was produced by immunizing rabbits with red blood cells from a rhesus macaque. The antigen that induced this immunization was designated by them as Rh factor to indicate that rhesus blood had been used for the production of the serum.[49]
In 1939, Phillip Levine and Rufus Stetson published in a first case report the clinical consequences of non-recognized Rh factor, hemolytic transfusion reaction, and hemolytic disease of the newborn in its most severe form.[50] It was recognized that the serum of the reported woman agglutinated with red blood cells of about 80% of the people although the then known blood groups, in particular ABO were matched. No name was given to this agglutinin when described. In 1940, Landsteiner and Wiener made the connection to their earlier discovery, reporting a serum that also reacted with about 85% of different human red blood cells.[51]
In 1941, Group O: a patient in Irvington, New Jersey, US, delivered a normal[clarification needed] infant in 1931; this pregnancy was followed by a long period of sterility. The second pregnancy (April, 1941) resulted in an infant with icterus gravis.[52] In May 1941, the third anti-Rh serum (M.S.) of Group O became available.[52]
Based on the serologic similarities, 'Rh factor' was later also used for antigens, and anti-Rh for antibodies, found in humans such as those previously described by Levine and Stetson. Although differences between these two sera were shown already in 1942 and clearly demonstrated in 1963, the already widely used term "Rh" was kept for the clinically described human antibodies which are different from the ones related to the rhesus monkey. This real factor found in rhesus macaque was classified in the Landsteiner-Weiner antigen system (antigen LW, antibody anti-LW) in honor of the discoverers.[53][54]
It was recognized that the Rh factor was just one in a system of various antigens. Based on different models of genetic inheritance, two different terminologies were developed; both of them are still in use.
The clinical significance of this highly immunizing D antigen (i.e., Rh factor) was soon realized. Some keystones were to recognize its importance for blood transfusion (including reliable diagnostic tests), hemolytic disease of the newborn (including exchange transfusion), and very importantly the prevention of it by screening and prophylaxis.
The discovery of cell-free fetal DNA in maternal circulation by Holzgrieve et al. led to the noninvasive genotyping of fetal Rh genes in many countries.
Related Research Articles
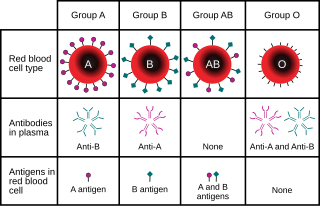
A blood type is a classification of blood, based on the presence and absence of antibodies and inherited antigenic substances on the surface of red blood cells (RBCs). These antigens may be proteins, carbohydrates, glycoproteins, or glycolipids, depending on the blood group system. Some of these antigens are also present on the surface of other types of cells of various tissues. Several of these red blood cell surface antigens can stem from one allele and collectively form a blood group system.
Rh disease is a type of hemolytic disease of the fetus and newborn (HDFN). HDFN due to anti-D antibodies is the proper and currently used name for this disease as the Rh blood group system actually has more than 50 antigens and not only the D-antigen. The term "Rh Disease" is commonly used to refer to HDFN due to anti-D antibodies, and prior to the discovery of anti-Rho(D) immune globulin, it was the most common type of HDFN. The disease ranges from mild to severe, and occurs in the second or subsequent pregnancies of Rh-D negative women when the biologic father is Rh-D positive.
Hemolytic disease of the newborn, also known as hemolytic disease of the fetus and newborn, HDN, HDFN, or erythroblastosis fetalis, is an alloimmune condition that develops in a fetus at or around birth, when the IgG molecules produced by the mother pass through the placenta. Among these antibodies are some which attack antigens on the red blood cells in the fetal circulation, breaking down and destroying the cells. The fetus can develop reticulocytosis and anemia. The intensity of this fetal disease ranges from mild to very severe, and fetal death from heart failure can occur. When the disease is moderate or severe, many erythroblasts are present in the fetal blood, earning these forms of the disease the name erythroblastosis fetalis.
The direct and indirect Coombs tests, also known as antiglobulin test (AGT), are blood tests used in immunohematology. The direct Coombs test detects antibodies that are stuck to the surface of the red blood cells. Since these antibodies sometimes destroy red blood cells they can cause anemia; this test can help clarify the condition. The indirect Coombs test detects antibodies that are floating freely in the blood. These antibodies could act against certain red blood cells; the test can be carried out to diagnose reactions to a blood transfusion.
Hemolytic disease of the newborn (anti-Kell1) is the second most common cause of severe hemolytic disease of the newborn (HDN) after Rh disease. Anti-Kell1 is becoming relatively more important as prevention of Rh disease is also becoming more effective.
Hemolytic disease of the newborn (anti-Rhc) can range from a mild to a severe disease. It is the third most common cause of severe HDN. Rh disease is the most common and hemolytic disease of the newborn (anti-Kell) is the second most common cause of severe HDN. It occurs more commonly in women who are Rh D negative.
The term human blood group systems is defined by the International Society of Blood Transfusion (ISBT) as systems in the human species where cell-surface antigens—in particular, those on blood cells—are "controlled at a single gene locus or by two or more very closely linked homologous genes with little or no observable recombination between them", and include the common ABO and Rh (Rhesus) antigen systems, as well as many others; 44 human systems are identified as of December 2022.
The Kell antigen system is a human blood group system, that is, a group of antigens on the human red blood cell surface which are important determinants of blood type and are targets for autoimmune or alloimmune diseases which destroy red blood cells. The Kell antigens are K, k, Kpa, Kpb, Jsa and Jsb. The Kell antigens are peptides found within the Kell protein, a 93-kilodalton transmembrane zinc-dependent endopeptidase which is responsible for cleaving endothelin-3.
The Kidd antigen system are proteins found in the Kidd's blood group, which act as antigens, i.e., they have the ability to produce antibodies under certain circumstances. The Jk antigen is found on a protein responsible for urea transport in the red blood cells and the kidney. They are important in transfusion medicine. People with two Jk(a) antigens, for instance, may form antibodies against donated blood containing two Jk(b) antigens. This can lead to hemolytic anemia, in which the body destroys the transfused blood, leading to low red blood cell counts. Another disease associated with the Jk antigen is hemolytic disease of the newborn, in which a pregnant woman's body creates antibodies against the blood of her fetus, leading to destruction of the fetal blood cells. Hemolytic disease of the newborn associated with Jk antibodies is typically mild, though fatal cases have been reported.
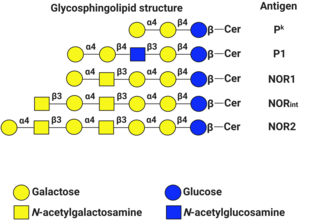
P1PK is a human blood group system based upon the A4GALT gene on chromosome 22. The P antigen was first described by Karl Landsteiner and Philip Levine in 1927. The P1PK blood group system consists of three glycosphingolipid antigens: Pk, P1 and NOR. In addition to glycosphingolipids, terminal Galα1→4Galβ structures are present on complex-type N-glycans. The GLOB antigen is now the member of the separate GLOB (globoside) blood group system.
Hemolytic disease of the newborn (anti-RhE) is caused by the anti-RhE antibody of the Rh blood group system. The anti-RhE antibody can be naturally occurring, or arise following immune sensitization after a blood transfusion or pregnancy.
Acquired hemolytic anemia can be divided into immune and non-immune mediated forms of hemolytic anemia.
Rh blood group, D antigen also known as Rh polypeptide 1 (RhPI) or cluster of differentiation 240D (CD240D) is a protein that in humans is encoded by the RHD gene.
Rh factor testing, also known as Rhesus factor testing, is the procedure of determining the Rhesus D status of an individual.
The Vel blood group is a human blood group that has been implicated in hemolytic transfusion reactions. The blood group consists of a single antigen, the high-frequency Vel antigen, which is expressed on the surface of red blood cells. Individuals are typed as Vel-positive or Vel-negative depending on the presence of this antigen. The expression of the antigen in Vel-positive individuals is highly variable and can range from strong to weak. Individuals with the rare Vel-negative blood type develop anti-Vel antibodies when exposed to Vel-positive blood, which can cause transfusion reactions on subsequent exposures.
The Junior blood group system is a human blood group defined by the presence or absence of the Jr(a) antigen, a high-frequency antigen that is found on the red blood cells of most individuals. People with the rare Jr(a) negative blood type can develop anti-Jr(a) antibodies, which may cause transfusion reactions and hemolytic disease of the newborn on subsequent exposures. Jr(a) negative blood is most common in people of Japanese heritage.
Blood compatibility testing is conducted in a medical laboratory to identify potential incompatibilities between blood group systems in blood transfusion. It is also used to diagnose and prevent some complications of pregnancy that can occur when the baby has a different blood group from the mother. Blood compatibility testing includes blood typing, which detects the antigens on red blood cells that determine a person's blood type; testing for unexpected antibodies against blood group antigens ; and, in the case of blood transfusions, mixing the recipient's plasma with the donor's red blood cells to detect incompatibilities (crossmatching). Routine blood typing involves determining the ABO and RhD type, and involves both identification of ABO antigens on red blood cells and identification of ABO antibodies in the plasma. Other blood group antigens may be tested for in specific clinical situations.
The Lan blood group system is a human blood group defined by the presence or absence of the Lan antigen on a person's red blood cells. More than 99.9% of people are positive for the Lan antigen. Individuals with the rare Lan-negative blood type, which is a recessive trait, can produce an anti-Lan antibody when exposed to Lan-positive blood. Anti-Lan antibodies may cause transfusion reactions on subsequent exposures to Lan-positive blood, and have also been implicated in mild cases of hemolytic disease of the newborn. However, the clinical significance of the antibody is variable. The antigen was first described in 1961, and Lan was officially designated a blood group in 2012.
The Sid blood group system is a human blood group defined by the presence or absence of the Sd(a) antigen on a person's red blood cells. About 96% of people are positive for the Sd(a) antigen, which is inherited as a dominant trait. Among Sd(a) positive individuals, the expression of the antigen ranges from extremely weak to extremely strong. Very strong expression of the antigen is referred to as a Sd(a++) phenotype. In addition to being expressed on red blood cells, Sd(a) is secreted in bodily fluids such as saliva and breast milk, and is found in the highest concentrations in urine. Urine testing is considered the most reliable method for determining a person's Sid blood type.
The Augustine blood group system is a human blood group system. It includes four red blood cell surface glycoprotein antigens which are encoded by alleles of the gene SLC29A1.
References
↑ Dean, Laura. Blood Groups and Red Cell Antigens [Internet].. Bethesda (MD): National Center for Biotechnology Information (US); 2005, Chapter. 7.
↑ Ottensooser F, Leon N, Sato M, Saldanha PH (March 1963). "Blood groups of a population of Ashkenazi Jews in Brazil". American Journal of Physical Anthropology. 21 (1): 41–8. doi:10.1002/ajpa.1330210106. PMID13940710. S2CID21795826.
↑ Harrison GA, Küchemann CF, Moore MA, Boyce AJ, Baju T, Mourant AE, etal. (1969). "The effects of altitudinal variation in Ethiopian populations". Philosophical Transactions of the Royal Society of London B: Biological Sciences. 256 (805): 147–182. Bibcode:1969RSPTB.256..147H. doi:10.1098/rstb.1969.0040.
↑ Touinssi M, Chiaroni J, Degioanni A, De Micco P, Dutour O, Bauduer F (2004). "Distribution of rhesus blood group system in the French basques: a reappraisal using the allele-specific primers PCR method". Human Heredity. 58 (2): 69–72. doi:10.1159/000083027. PMID15711086. S2CID44542508.
↑ "ABO inheritance patterns". Inheritance patterns of blood groups. Australian Red Cross Blood Service. Archived from the original on 1 November 2013. Retrieved 30 October 2013.
↑ Kustu S, Inwood W (2006). "Biological gas channels for NH3 and CO2: evidence that Rh (Rhesus) proteins are CO2 channels". Transfusion Clinique et Biologique. 13 (1–2): 103–10. doi:10.1016/j.tracli.2006.03.001. PMID16563833.
↑ Biver S, Scohy S, Szpirer J, Szpirer C, André B, Marini AM (2006). "Physiological role of the putative ammonium transporter RhCG in the mouse". Transfusion Clinique et Biologique. 13 (1–2): 167–8. doi:10.1016/j.tracli.2006.03.003. PMID16564721.
↑ Novotná M, Havlícek J, Smith AP, Kolbeková P, Skallová A, Klose J, etal. (September 2008). "Toxoplasma and reaction time: role of toxoplasmosis in the origin, preservation and geographical distribution of Rh blood group polymorphism". Parasitology. 135 (11): 1253–61. doi:10.1017/S003118200800485X. PMID18752708. S2CID5956901.
↑ Li CC (1953). "Is the Rh facing a crossroad? A critique of the compensation effect". Am Nat. 87 (835): 257–261. doi:10.1086/281782. S2CID84715943.
1 2 3 4 5 Brecher ME (2005). Technical Manual (15thed.). Bethesda MD: American Association of Blood Banks. ISBN978-1-56395-196-1.
1 2 3 Cartron JP (December 1999). "RH blood group system and molecular basis of Rh-deficiency". Baillière's Best Practice & Research. Clinical Haematology. 12 (4): 655–89. doi:10.1053/beha.1999.0047. PMID10895258.
↑ Landsteiner K, Wiener AS (1940). "An Agglutinable Factor in Human Blood Recognized by Immune Sera for Rhesus Blood". Exp Biol Med (Maywood). 43 (1): 223. doi:10.3181/00379727-43-11151. S2CID58298368.
↑ Landsteiner K, Wiener AS (1940). "An agglutinable factor in human blood recognized by immune sera for rhesus blood". Proc Soc Exp Biol Med. 43: 223–4. doi:10.3181/00379727-43-11151. S2CID58298368.
1 2 Levine P, Burnham L, Katzin E, Vogel P (December 1941). "The role of iso-immunization in the pathogenesis of erythroblastosis fetalis". American Journal of Obstetrics and Gynecology. 42 (6): 925–937. doi:10.1016/S0002-9378(41)90260-0. ISSN0002-9378.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.