
Homo habilis is an extinct species of archaic human from the Early Pleistocene of East and South Africa about 2.31 million years ago to 1.65 million years ago (mya). Upon species description in 1964, H. habilis was highly contested, with many researchers recommending it be synonymised with Australopithecus africanus, the only other early hominin known at the time, but H. habilis received more recognition as time went on and more relevant discoveries were made. By the 1980s, H. habilis was proposed to have been a human ancestor, directly evolving into Homo erectus which directly led to modern humans. This viewpoint is now debated. Several specimens with insecure species identification were assigned to H. habilis, leading to arguments for splitting, namely into "H. rudolfensis" and "H. gautengensis" of which only the former has received wide support.
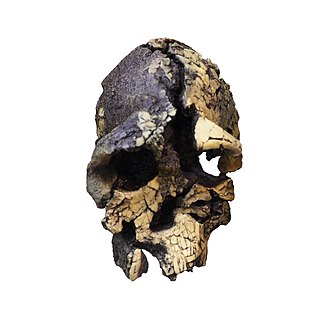
Kenyanthropus is a hominin genus identified from the Lomekwi site by Lake Turkana, Kenya, dated to 3.3 to 3.2 million years ago during the Middle Pliocene. It contains one species, K. platyops, but may also include the 2 million year old Homo rudolfensis, or K. rudolfensis. Before its naming in 2001, Australopithecus afarensis was widely regarded as the only australopithecine to exist during the Middle Pliocene, but Kenyanthropus evinces a greater diversity than once acknowledged. Kenyanthropus is most recognisable by an unusually flat face and small teeth for such an early hominin, with values on the extremes or beyond the range of variation for australopithecines in regard to these features. Multiple australopithecine species may have coexisted by foraging for different food items, which may be reason why these apes anatomically differ in features related to chewing.

Paranthropus is a genus of extinct hominin which contains two widely accepted species: P. robustus and P. boisei. However, the validity of Paranthropus is contested, and it is sometimes considered to be synonymous with Australopithecus. They are also referred to as the robust australopithecines. They lived between approximately 2.9 and 1.2 million years ago (mya) from the end of the Pliocene to the Middle Pleistocene.

Homo ergaster is an extinct species or subspecies of archaic humans who lived in Africa in the Early Pleistocene. Whether H. ergaster constitutes a species of its own or should be subsumed into H. erectus is an ongoing and unresolved dispute within palaeoanthropology. Proponents of synonymisation typically designate H. ergaster as "African Homo erectus" or "Homo erectus ergaster". The name Homo ergaster roughly translates to "working man", a reference to the more advanced tools used by the species in comparison to those of their ancestors. The fossil range of H. ergaster mainly covers the period of 1.7 to 1.4 million years ago, though a broader time range is possible. Though fossils are known from across East and Southern Africa, most H. ergaster fossils have been found along the shores of Lake Turkana in Kenya. There are later African fossils, some younger than 1 million years ago, that indicate long-term anatomical continuity, though it is unclear if they can be formally regarded as H. ergaster specimens. As a chronospecies, H. ergaster may have persisted to as late as 600,000 years ago, when new lineages of Homo arose in Africa.

Homo is the genus that emerged in the genus Australopithecus that encompasses the extant species Homo sapiens, along with several extinct species classified as either ancestral to or closely related to modern humans, including Homo erectus and Homo neanderthalensis. The oldest member of Homo is Homo habilis with records of just over 2 million years ago. Homo, together with the genus Paranthropus, is probably sister to Australopithecus africanus, which itself had previously split from the lineage of Pan, the chimpanzees.
Paleoanthropology or paleo-anthropology is a branch of paleontology and anthropology which seeks to understand the early development of anatomically modern humans, a process known as hominization, through the reconstruction of evolutionary kinship lines within the family Hominidae, working from biological evidence and cultural evidence.

Paranthropus aethiopicus is an extinct species of robust australopithecine from the Late Pliocene to Early Pleistocene of East Africa about 2.7–2.3 million years ago. However, it is much debated whether or not Paranthropus is an invalid grouping and is synonymous with Australopithecus, so the species is also often classified as Australopithecus aethiopicus. Whatever the case, it is considered to have been the ancestor of the much more robust P. boisei. It is debated if P. aethiopicus should be subsumed under P. boisei, and the terms P. boisei sensu lato and P. boisei sensu stricto can be used to respectively include and exclude P. aethiopicus from P. boisei.

Australopithecus garhi is a species of australopithecine from the Bouri Formation in the Afar Region of Ethiopia 2.6–2.5 million years ago (mya) during the Early Pleistocene. The first remains were described in 1999 based on several skeletal elements uncovered in the three years preceding. A. garhi was originally considered to have been a direct ancestor to Homo and the human line, but is now thought to have been an offshoot. Like other australopithecines, A. garhi had a brain volume of 450 cc (27 cu in); a jaw which jutted out (prognathism); relatively large molars and premolars; adaptations for both walking on two legs (bipedalism) and grasping while climbing (arboreality); and it is possible that, though unclear if, males were larger than females. One individual, presumed female based on size, may have been 140 cm tall.

Paranthropus robustus is a species of robust australopithecine from the Early and possibly Middle Pleistocene of the Cradle of Humankind, South Africa, about 2.27 to 0.87 million years ago. It has been identified in Kromdraai, Swartkrans, Sterkfontein, Gondolin, Cooper's, and Drimolen Caves. Discovered in 1938, it was among the first early hominins described, and became the type species for the genus Paranthropus. However, it has been argued by some that Paranthropus is an invalid grouping and synonymous with Australopithecus, so the species is also often classified as Australopithecus robustus.

Paranthropus boisei is a species of australopithecine from the Early Pleistocene of East Africa about 2.5 to 1.15 million years ago. The holotype specimen, OH 5, was discovered by palaeoanthropologist Mary Leakey in 1959, and described by her husband Louis a month later. It was originally placed into its own genus as "Zinjanthropus boisei", but is now relegated to Paranthropus along with other robust australopithecines. However, it is also argued that Paranthropus is an invalid grouping and synonymous with Australopithecus, so the species is also often classified as Australopithecus boisei.
Human taxonomy is the classification of the human species within zoological taxonomy. The systematic genus, Homo, is designed to include both anatomically modern humans and extinct varieties of archaic humans. Current humans have been designated as subspecies Homo sapiens sapiens, differentiated, according to some, from the direct ancestor, Homo sapiens idaltu.

Koobi Fora refers primarily to a region around Koobi Fora Ridge, located on the eastern shore of Lake Turkana in the territory of the nomadic Gabbra people. According to the National Museums of Kenya, the name comes from the Gabbra language:
In the language of the Gabbra people who live near the site, the term Koobi Fora means a place of the commiphora and the source of myrrh...

KNM ER 1813 is a skull of the species Homo habilis. It was discovered in Koobi Fora, Kenya by Kamoya Kimeu in 1973, and is estimated to be 1.9 million years old.
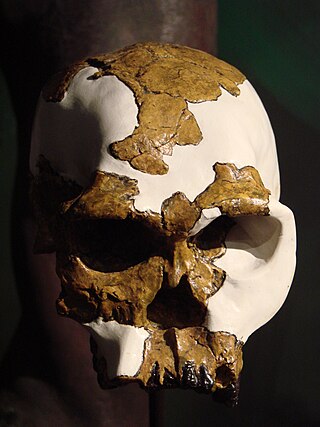
OH 24 is a fossilized skull of the species Homo habilis. It was discovered in Olduvai Gorge, Tanzania by Peter Nzube in 1968. The skull was found crushed almost flat and was therefore named after the famously skinny model of the time Twiggy. Estimated at 1.8 mya, the cranium was found crushed flat and cemented together with a mass coating of limestone. It is now a Smithsonian exhibit item.
KNM ER 406 is an almost complete fossilized skull of the species Paranthropus boisei. It was discovered in Koobi Fora, Kenya by Richard Leakey and H. Mutua in 1969. This species is grouped with the Australopitecine genus, Paranthropus boisei because of the robusticity of the skull and the prominent characteristics. This species was found well preserved with a complete cranium but lacking dentition. He was known for his robust cranial features that showed the signs of adaptation of the ecological niches. The big chewing muscles attached to the sagittal crest are traits of this adaptation.

Homo erectus is an extinct species of archaic human from the Pleistocene, with its earliest occurrence about 2 million years ago. Its specimens are among the first recognizable members of the genus Homo.

Australopithecus sediba is an extinct species of australopithecine recovered from Malapa Cave, Cradle of Humankind, South Africa. It is known from a partial juvenile skeleton, the holotype MH1, and a partial adult female skeleton, the paratype MH2. They date to about 1.98 million years ago in the Early Pleistocene, and coexisted with Paranthropus robustus and Homo ergaster / Homo erectus. Malapa is interpreted as having been a natural death trap, the base of a long vertical shaft which creatures could accidentally fall into. A. sediba was initially described as being a potential human ancestor, and perhaps the progenitor of Homo, but this is contested and it could also represent a late-surviving population or sister species of A. africanus which had earlier inhabited the area.
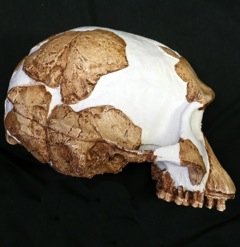
Homo gautengensis is a species name proposed by anthropologist Darren Curnoe in 2010 for South African hominin fossils otherwise attributed to H. habilis, H. ergaster, or, in some cases, Australopithecus or Paranthropus. The fossils assigned to the species by Curnoe cover a vast temporal range, from about 1.8 million years ago to potentially as late as 0.8 million years ago, meaning that if the species is considered valid, H. gautengensis would be both one of the earliest and one of the longest lived species of Homo.
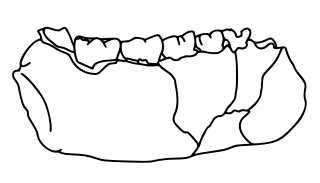
LD 350-1 is the earliest known specimen of the genus Homo, dating to 2.8–2.75 million years ago (mya), found in the Ledi-Geraru site in the Afar Region of Ethiopia. The specimen was discovered in silts 10 m (33 ft) above the Gurumaha Tuff section of the site by Ethiopian palaeoanthropologist Chalachew Seyoum on 29 January 2013. It is currently unassigned to a species, and it is unclear if it represents the ancestor to H. habilis and H. rudolfensis which evolved around 2.4 mya.
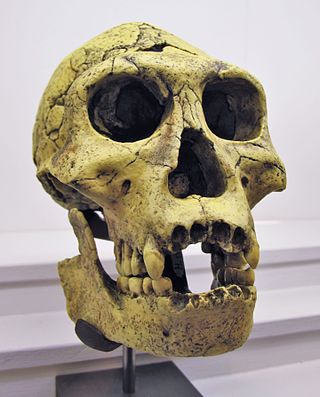
The Dmanisi hominins, Dmanisi people, or Dmanisi man were a population of Early Pleistocene hominins whose fossils have been recovered at Dmanisi, Georgia. The fossils and stone tools recovered at Dmanisi range in age from 1.85–1.77 million years old, making the Dmanisi hominins the earliest well-dated hominin fossils in Eurasia and the best preserved fossils of early Homo from a single site so early in time, though earlier fossils and artifacts have been found in Asia. Though their precise classification is controversial and disputed, the Dmanisi fossils are highly significant within research on early hominin migrations out of Africa. The Dmanisi hominins are known from over a hundred postcranial fossils and five famous well-preserved skulls, referred to as Dmanisi Skulls 1–5.





