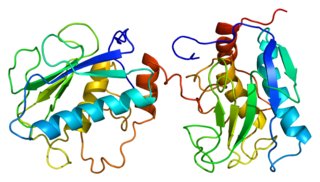Structure
The MMPs have a common domain structure. The three common domains are the pro-peptide, the catalytic domain, and the haemopexin-like C-terminal domain, which is linked to the catalytic domain by a flexible hinge region. [2]
The pro-peptide
The MMPs are initially synthesized as inactive zymogens with a pro-peptide domain that must be removed before the enzyme is active. The pro-peptide domain is part of the "cysteine switch." This contains a conserved cysteine residue that interacts with the zinc in the active site and prevents binding and cleavage of the substrate, keeping the enzyme in an inactive form. In the majority of the MMPs, the cysteine residue is in the conserved sequence PRCGxPD. Some MMPs have a prohormone convertase cleavage site (Furin-like) as part of this domain, which, when cleaved, activates the enzyme. MMP-23A and MMP-23B include a transmembrane segment in this domain. [9]
The catalytic domain
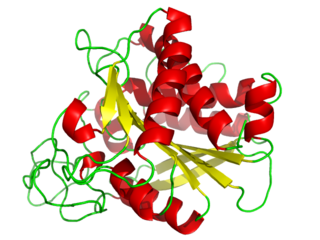
X-ray crystallographic structures of several MMP catalytic domains have shown that this domain is an oblate sphere measuring 35 x 30 x 30 Å (3.5 × 3 x 3 nm). The active site is a 20 Å (2 nm) groove that runs across the catalytic domain. In the part of the catalytic domain forming the active site there is a catalytically important Zn 2+ ion, which is bound by three histidine residues found in the conserved sequence HExxHxxGxxH. Hence, this sequence is a zinc-binding motif.
The gelatinases, such as MMP-2, incorporate Fibronectin type II modules inserted immediately before in the zinc-binding motif in the catalytic domain. [10]
The hinge region
The catalytic domain is connected to the C-terminal domain by a flexible hinge or linker region. This is up to 75 amino acids long, and has no determinable structure.
The hemopexin-like C-terminal domain

The C-terminal domain has structural similarities to the serum protein hemopexin. It has a four-bladed β-propeller structure. β-Propeller structures provide a large flat surface that is thought to be involved in protein-protein interactions. This determines substrate specificity and is the site for interaction with TIMP's (tissue inhibitor of metalloproteinases). The hemopexin-like domain is absent in MMP-7, MMP-23, MMP-26, and the plant and nematode. The membrane-bound MMPs (MT-MMPs) are anchored to the plasma membrane via a transmembrane or a GPI-anchoring domain.