Related Research Articles

In multicellular organisms, stem cells are undifferentiated or partially differentiated cells that can change into various types of cells and proliferate indefinitely to produce more of the same stem cell. They are the earliest type of cell in a cell lineage. They are found in both embryonic and adult organisms, but they have slightly different properties in each. They are usually distinguished from progenitor cells, which cannot divide indefinitely, and precursor or blast cells, which are usually committed to differentiating into one cell type.

Bone marrow is a semi-solid tissue found within the spongy portions of bones. In birds and mammals, bone marrow is the primary site of new blood cell production. It is composed of hematopoietic cells, marrow adipose tissue, and supportive stromal cells. In adult humans, bone marrow is primarily located in the ribs, vertebrae, sternum, and bones of the pelvis. Bone marrow comprises approximately 5% of total body mass in healthy adult humans, such that a man weighing 73 kg (161 lbs) will have around 3.7 kg (8 lbs) of bone marrow.
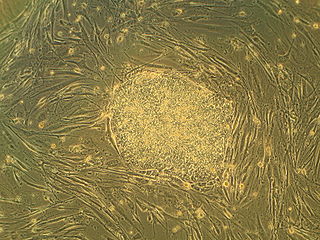
Embryonic stem cells (ESCs) are pluripotent stem cells derived from the inner cell mass of a blastocyst, an early-stage pre-implantation embryo. Human embryos reach the blastocyst stage 4–5 days post fertilization, at which time they consist of 50–150 cells. Isolating the inner cell mass (embryoblast) using immunosurgery results in destruction of the blastocyst, a process which raises ethical issues, including whether or not embryos at the pre-implantation stage have the same moral considerations as embryos in the post-implantation stage of development.

Cell therapy is a therapy in which viable cells are injected, grafted or implanted into a patient in order to effectuate a medicinal effect, for example, by transplanting T-cells capable of fighting cancer cells via cell-mediated immunity in the course of immunotherapy, or grafting stem cells to regenerate diseased tissues.
Stromal cells, or mesenchymal stromal cells, are differentiating cells found in abundance within bone marrow but can also be seen all around the body. Stromal cells can become connective tissue cells of any organ, for example in the uterine mucosa (endometrium), prostate, bone marrow, lymph node and the ovary. They are cells that support the function of the parenchymal cells of that organ. The most common stromal cells include fibroblasts and pericytes. The term stromal comes from Latin stromat-, "bed covering", and Ancient Greek στρῶμα, strôma, "bed".
Adult stem cells are undifferentiated cells, found throughout the body after development, that multiply by cell division to replenish dying cells and regenerate damaged tissues. Also known as somatic stem cells, they can be found in juvenile, adult animals, and humans, unlike embryonic stem cells.

A stem cell line is a group of stem cells that is cultured in vitro and can be propagated indefinitely. Stem cell lines are derived from either animal or human tissues and come from one of three sources: embryonic stem cells, adult stem cells, or induced stem cells. They are commonly used in research and regenerative medicine.
Stem-cell therapy uses stem cells to treat or prevent a disease or condition. As of 2016, the only established therapy using stem cells is hematopoietic stem cell transplantation. This usually takes the form of a bone marrow transplantation, but the cells can also be derived from umbilical cord blood. Research is underway to develop various sources for stem cells as well as to apply stem-cell treatments for neurodegenerative diseases and conditions such as diabetes and heart disease.

Induced pluripotent stem cells are a type of pluripotent stem cell that can be generated directly from a somatic cell. The iPSC technology was pioneered by Shinya Yamanaka and Kazutoshi Takahashi in Kyoto, Japan, who together showed in 2006 that the introduction of four specific genes, collectively known as Yamanaka factors, encoding transcription factors could convert somatic cells into pluripotent stem cells. Shinya Yamanaka was awarded the 2012 Nobel Prize along with Sir John Gurdon "for the discovery that mature cells can be reprogrammed to become pluripotent."
Mesenchymal stem cells (MSCs) are multipotent cells found in multiple human adult tissues, including bone marrow, synovial tissues, and adipose tissues. Since they are derived from the mesoderm, they have been shown to differentiate into bone, cartilage, muscle, and adipose tissue. MSCs from embryonic sources have shown promise scientifically while creating significant controversy. As a result, many researchers have focused on adult stem cells, or stem cells isolated from adult humans that can be transplanted into damaged tissue.
Dental pulp stem cells (DPSCs) are stem cells present in the dental pulp, which is the soft living tissue within teeth. DPSCs can be collected from dental pulp by means of a non-invasive practice. It can be performed with an adult after simple extraction or to the young after surgical extraction of wisdom teeth. They are pluripotent, as they can form embryoid body-like structures (EBs) in vitro and teratoma-like structures that contained tissues derived from all three embryonic germ layers when injected in nude mice. DPSCs can differentiate in vitro into tissues that have similar characteristics to mesoderm, endoderm and ectoderm layers. They can differentiate into many cell types, such as odontoblasts, neural progenitors, osteoblasts, chondrocytes, and adipocytes. DPSCs were found to be able to differentiate into adipocytes and neural-like cells. DPSC differentiation into osteogenic lines is enhanced in 3D condition and hypoxia. These cells can be obtained from postnatal teeth, wisdom teeth, and deciduous teeth, providing researchers with a non-invasive method of extracting stem cells. The different cell populations, however, differ in certain aspects of their growth rate in culture, marker gene expression and cell differentiation, although the extent to which these differences can be attributed to tissue of origin, function or culture conditions remains unclear. As a result, DPSCs have been thought of as an extremely promising source of cells used in endogenous tissue engineering.

Cell potency is a cell's ability to differentiate into other cell types. The more cell types a cell can differentiate into, the greater its potency. Potency is also described as the gene activation potential within a cell, which like a continuum, begins with totipotency to designate a cell with the most differentiation potential, pluripotency, multipotency, oligopotency, and finally unipotency.
Mesenchymal stem cells (MSCs) also known as mesenchymal stromal cells or medicinal signaling cells, are multipotent stromal cells that can differentiate into a variety of cell types, including osteoblasts, chondrocytes, myocytes and adipocytes.
Adult mesenchymal stem cells are being used by researchers in the fields of regenerative medicine and tissue engineering to artificially reconstruct human tissue which has been previously damaged. Mesenchymal stem cells are able to differentiate, or mature from a less specialized cell to a more specialized cell type, to replace damaged tissues in various organs.
The Stem Cell Lineage Database (SCLD) is a database of resources used to identify cell lineages.
Stage-specific embryonic antigen 3 (SSEA-3) is a glycosphingolipid, specifically, an oligosaccharide composed of five carbohydrate units connected to a sphingolipid. Sphingolipids were originally discovered in 1884 by Johann Ludwig Wilhelm Thudichum who named them after the Sphinx of Greek mythology in reference to the unresolved riddle of their function. It is now known that sphingolipids function as key players in cell signaling and the SSEA-3 molecule as a whole plays a key role in identifying many types of mammalian cells with pluripotent and stem cell-like characteristics.
Dermal fibroblasts are cells within the dermis layer of skin which are responsible for generating connective tissue and allowing the skin to recover from injury. Using organelles, dermal fibroblasts generate and maintain the connective tissue which unites separate cell layers. Furthermore, these dermal fibroblasts produce the protein molecules including laminin and fibronectin which comprise the extracellular matrix. By creating the extracellular matrix between the dermis and epidermis, fibroblasts allow the epithelial cells of the epidermis to affix the matrix, thereby allowing the epidermal cells to effectively join together to form the top layer of the skin.
Induced stem cells (iSC) are stem cells derived from somatic, reproductive, pluripotent or other cell types by deliberate epigenetic reprogramming. They are classified as either totipotent (iTC), pluripotent (iPSC) or progenitor or unipotent – (iUSC) according to their developmental potential and degree of dedifferentiation. Progenitors are obtained by so-called direct reprogramming or directed differentiation and are also called induced somatic stem cells.
Cord lining, cord tissue, or umbilical cord lining membrane, is the outermost layer of the umbilical cord. As the umbilical cord itself is an extension of the placenta, the umbilical cord lining membrane is an extension of the amniotic membrane covering the placenta. The umbilical cord lining membrane comprises two layers: the amniotic layer and the sub-amniotic layer. The umbilical cord lining membrane is a rich source of two strains of stem cells (CLSCs): epithelial stem cells (CLECs) and mesenchymal stem cells (CLMCs). Discovered by Singapore-based CellResearch Corporation in 2004, this is the best known source for harvesting human stem cells.

Stem cellfat grafting is autotransplantation of adipose-derived stem cells (ADSCs) extracted from fat-abundant donor sites to other areas such as the face, breast, and hip to reconstruct the operative areas into desirable shapes. ADSCs are multipotent stem cells found in adipose tissues, displaying similar differentiation potentials to bone marrow-derived mesenchymal stem cells (BM-MSCs).
References
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 Kuroda, Y.; Kitada, M.; Wakao, S.; Nishikawa, K.; Tanimura, Y.; Makinoshima, H.; Goda, M.; Akashi, H.; Inutsuka, A.; Niwa, A.; Shigemoto, T.; Nabeshima, Y.; Nakahata, T.; Nabeshima, Y.-i.; Fujiyoshi, Y.; Dezawa, M. (2010). "Unique multipotent cells in adult human mesenchymal cell populations". Proceedings of the National Academy of Sciences. 107 (19): 8639–43. Bibcode:2010PNAS..107.8639K. doi: 10.1073/pnas.0911647107 . PMC 2889306 . PMID 20421459.
- 1 2 Dezawa, Mari, ed. (2018-11-27). Muse Cells: Endogenous Reparative Pluripotent Stem Cells. Advances in Experimental Medicine and Biology. Vol. 103. Springer Tokyo. doi:10.1007/978-4-431-56847-6. ISBN 978-4-431-56847-6.
- ↑ Leng, Zikuan; Sun, Dongming; Huang, Zihao; Tadmori, Iman; Chiang, Ning; Kethidi, Nikhit; Sabra, Ahmed; Kushida, Yoshihiro; Fu, Yu-Show; Dezawa, Mari; He, Xijing; Young, Wise (2019-04-18). "Quantitative Analysis of SSEA3+ Cells from Human Umbilical Cord after Magnetic Sorting". Cell Transplantation. 28 (7): 907–923. doi: 10.1177/0963689719844260 . PMC 6719495 . PMID 30997834. S2CID 122342163.
- 1 2 3 4 5 6 7 8 9 10 Wakao, S.; Kitada, M.; Kuroda, Y.; Shigemoto, T.; Matsuse, D.; Akashi, H.; Tanimura, Y.; Tsuchiyama, K.; Kikuchi, T.; Goda, M.; Nakahata, T.; Fujiyoshi, Y.; Dezawa, M. (2011). "Multilineage-differentiating stress-enduring (Muse) cells are a primary source of induced pluripotent stem cells in human fibroblasts". Proceedings of the National Academy of Sciences. 108 (24): 9875–80. Bibcode:2011PNAS..108.9875W. doi: 10.1073/pnas.1100816108 . PMC 3116385 . PMID 21628574.
- 1 2 3 4 Dezawa, Mari (2016). "Muse Cells Provide the Pluripotency of Mesenchymal Stem Cells: Direct Contribution of Muse Cells to Tissue Regeneration". Cell Transplantation. 25 (5): 849–61. doi: 10.3727/096368916X690881 . PMID 26884346.
- 1 2 3 Hori, Emiko; Hayakawa, Yumiko; Hayashi, Tomohide; Hori, Satoshi; Okamoto, Soushi; Shibata, Takashi; Kubo, Michiya; Horie, Yukio; Sasahara, Masakiyo; Kuroda, Satoshi (2016). "Mobilization of Pluripotent Multilineage-Differentiating Stress-Enduring Cells in Ischemic Stroke". Journal of Stroke and Cerebrovascular Diseases. 25 (6): 1473–81. doi:10.1016/j.jstrokecerebrovasdis.2015.12.033. PMID 27019988.
- 1 2 Kuroda, Yasumasa; Wakao, Shohei; Kitada, Masaaki; Murakami, Toru; Nojima, Makoto; Dezawa, Mari (2013). "Isolation, culture and evaluation of multilineage-differentiating stress-enduring (Muse) cells". Nature Protocols. 8 (7): 1391–415. doi:10.1038/nprot.2013.076. PMID 23787896. S2CID 28597290.[ unreliable medical source? ]
- 1 2 3 Ogura, Fumitaka; Wakao, Shohei; Kuroda, Yasumasa; Tsuchiyama, Kenichiro; Bagheri, Mozhdeh; Heneidi, Saleh; Chazenbalk, Gregorio; Aiba, Setsuya; Dezawa, Mari (2014). "Human Adipose Tissue Possesses a Unique Population of Pluripotent Stem Cells with Nontumorigenic and Low Telomerase Activities: Potential Implications in Regenerative Medicine". Stem Cells and Development. 23 (7): 717–28. doi:10.1089/scd.2013.0473. PMID 24256547.
- 1 2 3 Heneidi, Saleh; Simerman, Ariel A.; Keller, Erica; Singh, Prapti; Li, Xinmin; Dumesic, Daniel A.; Chazenbalk, Gregorio (2013). "Awakened by Cellular Stress: Isolation and Characterization of a Novel Population of Pluripotent Stem Cells Derived from Human Adipose Tissue". PLOS ONE. 8 (6): e64752. Bibcode:2013PLoSO...864752H. doi: 10.1371/journal.pone.0064752 . PMC 3673968 . PMID 23755141.
- 1 2 3 Noda, Toshiyuki; Nishigaki, Kazuhiko; Minatoguchi, Shinya (2020-06-25). "Safety and Efficacy of Human Muse Cell-Based Product for Acute Myocardial Infarction in a First-in-Human Trial". Circulation Journal: Official Journal of the Japanese Circulation Society. 84 (7): 1189–1192. doi: 10.1253/circj.CJ-20-0307 . ISSN 1347-4820. PMID 32522904.
- 1 2 3 4 Niizuma, Kuniyasu; Osawa, Shin-Ichiro; Endo, Hidenori; Izumi, Shin-Ichi; Ataka, Kota; Hirakawa, Akihiro; Iwano, Masao; Tominaga, Teiji (December 2023). "Randomized placebo-controlled trial of CL2020, an allogenic muse cell-based product, in subacute ischemic stroke". Journal of Cerebral Blood Flow and Metabolism: Official Journal of the International Society of Cerebral Blood Flow and Metabolism. 43 (12): 2029–2039. doi:10.1177/0271678X231202594. ISSN 1559-7016. PMC 10925866 . PMID 37756573.
- 1 2 3 Fujita, Y.; Nohara, T.; Takashima, S.; Natsuga, K.; Adachi, M.; Yoshida, K.; Shinkuma, S.; Takeichi, T.; Nakamura, H.; Wada, O.; Akiyama, M.; Ishiko, A.; Shimizu, H. (August 2021). "Intravenous allogeneic multilineage-differentiating stress-enduring cells in adults with dystrophic epidermolysis bullosa: a phase 1/2 open-label study". Journal of the European Academy of Dermatology and Venereology: JEADV. 35 (8): e528–e531. doi:10.1111/jdv.17201. ISSN 1468-3083. PMC 8359848 . PMID 33656198.
- 1 2 3 Yamashita, Toru; Nakano, Yumiko; Sasaki, Ryo; Tadokoro, Koh; Omote, Yoshio; Yunoki, Taijun; Kawahara, Yuko; Matsumoto, Namiko; Taira, Yuki; Matsuoka, Chika; Morihara, Ryuta; Abe, Koji (2023). "Safety and Clinical Effects of a Muse Cell-Based Product in Patients With Amyotrophic Lateral Sclerosis: Results of a Phase 2 Clinical Trial". Cell Transplantation. 32: 9636897231214370. doi:10.1177/09636897231214370. ISSN 1555-3892. PMC 10686030 . PMID 38014622.
- ↑ "Muse細胞製品を用いた新生児低酸素性虚血性脳症に対する 医師主導治験開始に関するお知らせについて".
- ↑ https://www.lsii.co.jp/assets/pdf/en/20210518-1.pdf [ bare URL PDF ]
- ↑ Alessio, Nicola; Özcan, Servet; Tatsumi, Kazuki; Murat, Ayşegül; Peluso, Gianfranco; Dezawa, Mari; Galderisi, Umberto (2016). "The secretome of MUSE cells contains factors that may play a role in regulation of stemness, apoptosis and immunomodulation". Cell Cycle. 16 (1): 1–12. doi:10.1080/15384101.2016.1211215. PMC 5270533 . PMID 27463232.
- ↑ Wakao, S; Kitada, M; Kuroda, Y; Shigemoto, T; Matsuse, D; Akashi, H; Tanimura, Y; Tsuchiyama, K; Kikuchi, T; Goda, M; Nakahata, T; Fujiyoshi, Y; Dezawa, M (2011). "Multilineage-differentiating stress-enduring (Muse) cells are a primary source of induced pluripotent stem cells in human fibroblasts". Proceedings of the National Academy of Sciences. 108 (24): 9875–9880. Bibcode:2011PNAS..108.9875W. doi: 10.1073/pnas.1100816108 . PMC 3116385 . PMID 21628574.
- ↑ Alessio, N; Squillaro, T; Özcan, S; Di Bernardo, G; Venditti, M; Melone, M; Peluso, G; Galderisi, U (2018). "Stress and stem cells: Adult Muse cells tolerate extensive genotoxic stimuli better than mesenchymal stromal cells". Oncotarget. 9 (27): 19328–19341. doi:10.18632/oncotarget.25039. PMC 5922400 . PMID 29721206.
- 1 2 3 Gimeno, M. L.; Fuertes, F.; Barcala Tabarrozzi, A. E.; Attorressi, A. I.; Cucchiani, R.; Corrales, L.; Oliveira, T. C.; Sogayar, M. C.; Labriola, L.; Dewey, R. A.; Perone, M. J. (2016). "Pluripotent Nontumorigenic Adipose Tissue-Derived Muse Cells Have Immunomodulatory Capacity Mediated by Transforming Growth Factor-β1". Stem Cells Translational Medicine. 6 (1): 161–173. doi:10.5966/sctm.2016-0014. PMC 5442729 . PMID 28170177.
- 1 2 3 4 Yamada, Y; Wakao, S; Kushida, Y; Minatoguchi, S; Mikami, A; Higashi, K; Baba, S; Shigemoto, T; Kuroda, Y; Kanamori, H; Amin, M; Kawasaki, M; Nishigaki, K; Taoka, M; Isobe, T; Muramatsu, C; Dezawa, M; Minayoguchi, S (2018-04-13). "S1P-S1PR2 Axis Mediates Homing of Muse Cells Into Damaged Heart for Long-Lasting Tissue Repair and Functional Recovery After Acute Myocardial Infarction". Ciruclation Research. 122 (8): 1069–1083. doi: 10.1161/CIRCRESAHA.117.311648 . PMID 29475983. S2CID 3503514.
- 1 2 3 4 Katagiri, H.; Kushida, Y.; Nojima, M.; Kuroda, Y.; Wakao, S.; Ishida, K.; Endo, F.; Kume, K.; Takahara, T.; Nitta, H.; Tsuda, H.; Dezawa, M.; Nishizuka, S. S. (2016). "A Distinct Subpopulation of Bone Marrow Mesenchymal Stem Cells, Muse Cells, Directly Commit to the Replacement of Liver Components". American Journal of Transplantation. 16 (2): 468–83. doi: 10.1111/ajt.13537 . PMID 26663569.
- 1 2 3 4 5 6 7 Uchida, Hiroki; Morita, Takahiro; Niizuma, Kuniyasu; Kushida, Yoshihiro; Kuroda, Yasumasa; Wakao, Shohei; Sakata, Hiroyuki; Matsuzaka, Yoshiya; Mushiake, Hajime; Tominaga, Teiji; Borlongan, Cesario V.; Dezawa, Mari (2016). "Transplantation of Unique Subpopulation of Fibroblasts, Muse Cells, Ameliorates Experimental Stroke Possibly via Robust Neuronal Differentiation". Stem Cells. 34 (1): 160–73. doi: 10.1002/stem.2206 . PMID 26388204.
- 1 2 3 4 5 Kinoshita, K.; Kuno, S.; Ishimine, H.; Aoi, N.; Mineda, K.; Kato, H.; Doi, K.; Kanayama, K.; Feng, J.; Mashiko, T.; Kurisaki, A.; Yoshimura, K. (2015). "Therapeutic Potential of Adipose-Derived SSEA-3-Positive Muse Cells for Treating Diabetic Skin Ulcers". Stem Cells Translational Medicine. 4 (2): 146–55. doi:10.5966/sctm.2014-0181. PMC 4303359 . PMID 25561682.
- 1 2 3 Kuroda, Yasumasa; Kitada, Masaaki; Wakao, Shohei; Nishikawa, Kouki; Tanimura, Yukihiro; Makinoshima, Hideki; Goda, Makoto; Akashi, Hideo; Inutsuka, Ayumu (2010-05-11). "Unique multipotent cells in adult human mesenchymal cell populations". Proceedings of the National Academy of Sciences of the United States of America. 107 (19): 8639–8643. Bibcode:2010PNAS..107.8639K. doi: 10.1073/pnas.0911647107 . ISSN 1091-6490. PMC 2889306 . PMID 20421459.
- ↑ Thomson, J. A.; Itskovitz-Eldor, Joseph; Shapiro, Sander S.; Waknitz, Michelle A.; Swiergiel, Jennifer J.; Marshall, Vivienne S.; Jones, Jeffrey M. (1998). "Embryonic Stem Cell Lines Derived from Human Blastocysts". Science. 282 (5391): 1145–7. Bibcode:1998Sci...282.1145T. doi: 10.1126/science.282.5391.1145 . PMID 9804556.
- 1 2 Wakao, Shohei; Kitada, Masaaki; Dezawa, Mari (2013). "The elite and stochastic model for iPS cell generation: Multilineage-differentiating stress enduring (Muse) cells are readily reprogrammable into iPS cells". Cytometry Part A. 83A (1): 18–26. doi: 10.1002/cyto.a.22069 . PMID 22693162.
- ↑ Tsuchiyama, Kenichiro; Wakao, Shohei; Kuroda, Yasumasa; Ogura, Fumitaka; Nojima, Makoto; Sawaya, Natsue; Yamasaki, Kenshi; Aiba, Setsuya; Dezawa, Mari (2013). "Functional Melanocytes Are Readily Reprogrammable from Multilineage-Differentiating Stress-Enduring (Muse) Cells, Distinct Stem Cells in Human Fibroblasts". Journal of Investigative Dermatology. 133 (10): 2425–35. doi: 10.1038/jid.2013.172 . PMID 23563197.
- 1 2 3 Wakao, Shohei; Oguma, Yo; Kushida, Yoshihiro; Kuroda, Yasumasa; Tatsumi, Kazuki; Dezawa, Mari (2022-10-06). "Phagocytosing differentiated cell-fragments is a novel mechanism for controlling somatic stem cell differentiation within a short time frame". Cellular and Molecular Life Sciences. 79 (11): 542. doi:10.1007/s00018-022-04555-0. ISSN 1420-9071. PMC 9537123 . PMID 36203068.
- 1 2 3 Fujita, Yasuyuki; Komatsu, Miho; Lee, San Eun; Kushida, Yoshihiro; Nakayama-Nishimura, Chihiro; Matsumura, Wakana; Takashima, Shota; Shinkuma, Satoru; Nomura, Toshifumi; Masutomi, Naoya; Kawamura, Makoto; Dezawa, Mari; Shimizu, Hiroshi (2020-06-12). "Intravenous Injection of Muse Cells as a Potential Therapeutic Approach for Epidermolysis Bullosa". Journal of Investigative Dermatology. 141 (1): 198–202. doi: 10.1016/j.jid.2020.05.092 . PMID 32540249. S2CID 219704432.
- 1 2 3 Mineda, K.; Feng, J.; Ishimine, H.; Takada, H.; Doi, K.; Kuno, S.; Kinoshita, K.; Kanayama, K.; Kato, H.; Mashiko, T.; Hashimoto, I.; Nakanishi, H.; Kurisaki, A.; Yoshimura, K. (2015). "Therapeutic Potential of Human Adipose-Derived Stem/Stromal Cell Microspheroids Prepared by Three-Dimensional Culture in Non-Cross-Linked Hyaluronic Acid Gel". Stem Cells Translational Medicine. 4 (12): 1511–22. doi:10.5966/sctm.2015-0037. PMC 4675504 . PMID 26494781.
- 1 2 3 4 Wakao, Shohei; Kuroda, Yasumasa; Ogura, Fumitaka; Shigemoto, Taeko; Dezawa, Mari (2012). "Regenerative Effects of Mesenchymal Stem Cells: Contribution of Muse Cells, a Novel Pluripotent Stem Cell Type that Resides in Mesenchymal Cells". Cells. 1 (4): 1045–60. doi: 10.3390/cells1041045 . PMC 3901150 . PMID 24710542.
- ↑ Kitada, Masaaki; Wakao, Shohei; Dezawa, Mari (2012). "Muse cells and induced pluripotent stem cell: Implication of the elite model". Cellular and Molecular Life Sciences. 69 (22): 3739–50. doi:10.1007/s00018-012-0994-5. PMC 3478511 . PMID 22527723.
- ↑ Chou, Yu-Fen; Chen, Hsu-Hsin; Eijpe, Maureen; Yabuuchi, Akiko; Chenoweth, Joshua G.; Tesar, Paul; Lu, Jun; McKay, Ronald D.G.; Geijsen, Niels (2008). "The Growth Factor Environment Defines Distinct Pluripotent Ground States in Novel Blastocyst-Derived Stem Cells". Cell. 135 (3): 449–61. doi:10.1016/j.cell.2008.08.035. PMC 2767270 . PMID 18984157.
- 1 2 3 Takahashi, Yoshiharu; Kajitani, Takumi; Endo, Toshiki; Nakayashiki, Atsushi; Inoue, Tomoo; Niizuma, Kuniyasu; Tominaga, Teiji (2023-09-27). "Intravenous Administration of Human Muse Cells Ameliorates Deficits in a Rat Model of Subacute Spinal Cord Injury". International Journal of Molecular Sciences. 24 (19): 14603. doi: 10.3390/ijms241914603 . ISSN 1422-0067. PMC 10572998 . PMID 37834052.
- 1 2 3 Nagaoki, Toshihide; Kumagai, Gentaro; Nitobe, Yohshiro; Sasaki, Ayako; Fujita, Taku; Fukutoku, Tatsuhiro; Saruta, Kenya; Tsukuda, Manami; Asari, Toru; Wada, Kanichiro; Dezawa, Mari; Ishibashi, Yasuyuki (December 2023). "Comparison of the Anti-Inflammatory Effects of Mouse Adipose- and Bone-Marrow-Derived Multilineage-Differentiating Stress-Enduring Cells in Acute-Phase Spinal Cord Injury". Journal of Neurotrauma. 40 (23–24): 2596–2609. doi:10.1089/neu.2022.0470. ISSN 1557-9042. PMID 37051701.
- 1 2 3 Kajitani, Takumi; Endo, Toshiki; Iwabuchi, Naoya; Inoue, Tomoo; Takahashi, Yoshiharu; Abe, Takatsugu; Niizuma, Kuniyasu; Tominaga, Teiji (2021-01-01). "Association of intravenous administration of human Muse cells with deficit amelioration in a rat model of spinal cord injury". Journal of Neurosurgery. Spine. 34 (4): 648–655. doi:10.3171/2020.7.SPINE20293. ISSN 1547-5646. PMID 33385996.
- 1 2 Abe, Takatsugu; Aburakawa, Daiki; Niizuma, Kuniyasu; Iwabuchi, Naoya; Kajitani, Takumi; Wakao, Shohei; Kushida, Yoshihiro; Dezawa, Mari; Borlongan, Cesar V.; Tominaga, Teiji (February 2020). "Intravenously Transplanted Human Multilineage-Differentiating Stress-Enduring Cells Afford Brain Repair in a Mouse Lacunar Stroke Model". Stroke. 51 (2): 601–611. doi:10.1161/STROKEAHA.119.026589. ISSN 1524-4628. PMID 31826733.
- 1 2 3 Uchida, Nao; Kushida, Yoshihiro; Kitada, Masaaki; Wakao, Shohei; Kumagai, Naonori; Kuroda, Yasumasa; Kondo, Yoshiaki; Hirohara, Yukari; Kure, Shigeo (2017-07-03). "Beneficial Effects of Systemically Administered Human Muse Cells in Adriamycin Nephropathy". Journal of the American Society of Nephrology. 28 (10): 2946–2960. doi:10.1681/ASN.2016070775. ISSN 1533-3450. PMC 5619953 . PMID 28674043.
- 1 2 Guo, Yonglong; Xue, Yunxia; Wang, Peiyuan; Cui, Zekai; Cao, Jixing; Liu, Shiwei; Yu, Quan; Zeng, Qiaolang; Zhu, Deliang; Xie, Mengyuan; Zhang, Jun (2020-09-23). "Muse cell spheroids have therapeutic effect on corneal scarring wound in mice and tree shrews". Science Translational Medicine. 12 (562): eaaw1120. doi:10.1126/scitranslmed.aaw1120. ISSN 1946-6242. PMID 32967971. S2CID 221860257.
- ↑ Nitobe, Yohshiro; Nagaoki, Toshihide; Kumagai, Gentaro; Sasaki, Ayako; Liu, Xizhe; Fujita, Taku; Fukutoku, Tatsuhiro; Wada, Kanichiro; Tanaka, Toshihiro; Kudo, Hitoshi; Asari, Toru; Furukawa, Ken-Ichi; Ishibashi, Yasuyuki (2019). "Neurotrophic Factor Secretion and Neural Differentiation Potential of Multilineage-differentiating Stress-enduring (Muse) Cells Derived from Mouse Adipose Tissue". Cell Transplantation. 28 (9–10): 1132–1139. doi:10.1177/0963689719863809. ISSN 1555-3892. PMC 6767880 . PMID 31304790.
- ↑ Liu, Jun; Yang, Zhongcai; Qiu, Mingning; Luo, Yan; Pang, Meijun; Wu, Yongyan; Zhang, Yong (April 2013). "Developmental potential of cloned goat embryos from an SSEA3(+) subpopulation of skin fibroblasts". Cellular Reprogramming. 15 (2): 159–165. doi:10.1089/cell.2012.0073. ISSN 2152-4998. PMID 23441574.
- 1 2 Iseki, Masahiro; Mizuma, Masamichi; Wakao, Shohei; Kushida, Yoshihiro; Kudo, Katsuyoshi; Fukase, Masahiko; Ishida, Masaharu; Ono, Tomoyuki; Shimura, Mitsuhiro; Ise, Ichiro; Suzuki, Yukie; Sueta, Teruko; Asada, Ryuta; Shimizu, Shinobu; Ueno, Yoshiyuki (April 2021). "The evaluation of the safety and efficacy of intravenously administered allogeneic multilineage-differentiating stress-enduring cells in a swine hepatectomy model". Surgery Today. 51 (4): 634–650. doi:10.1007/s00595-020-02117-0. ISSN 1436-2813. PMID 32915286.
- ↑ Mitani, Kosuke; Ito, Yuki; Takene, Yukio; Hatoya, Shingo; Sugiura, Kikuya; Inaba, Toshio (March 2021). "Long-Term Trypsin Treatment Promotes Stem Cell Potency of Canine Adipose-Derived Mesenchymal Stem Cells". Stem Cells and Development. 30 (6): 337–349. doi:10.1089/scd.2020.0175. ISSN 1557-8534. PMID 33528297.
- ↑ Byrne, James A.; Nguyen, Ha Nam; Reijo Pera, Renee A. (2009). "Enhanced Generation of Induced Pluripotent Stem Cells from a Subpopulation of Human Fibroblasts". PLOS ONE. 4 (9): e7118. Bibcode:2009PLoSO...4.7118B. doi: 10.1371/journal.pone.0007118 . PMC 2744017 . PMID 19774082.
- ↑ Tsuchiyama, Kenichiro; Wakao, Shohei; Kuroda, Yasumasa; Ogura, Fumitaka; Nojima, Makoto; Sawaya, Natsue; Yamasaki, Kenshi; Aiba, Setsuya; Dezawa, Mari (October 2013). "Functional melanocytes are readily reprogrammable from multilineage-differentiating stress-enduring (muse) cells, distinct stem cells in human fibroblasts". The Journal of Investigative Dermatology. 133 (10): 2425–2435. doi: 10.1038/jid.2013.172 . ISSN 1523-1747. PMID 23563197.
- ↑ Tian, Ting; Zhang, Ru-Zhi; Yang, Yu-Hua; Liu, Qi; Li, Di; Pan, Xiao-Ru (April 2017). "Muse Cells Derived from Dermal Tissues Can Differentiate into Melanocytes". Cellular Reprogramming. 19 (2): 116–122. doi:10.1089/cell.2016.0032. ISSN 2152-4998. PMID 28170296.
- ↑ Heneidi, Saleh; Simerman, Ariel A.; Keller, Erica; Singh, Prapti; Li, Xinmin; Dumesic, Daniel A.; Chazenbalk, Gregorio (2013). "Awakened by cellular stress: isolation and characterization of a novel population of pluripotent stem cells derived from human adipose tissue". PLOS ONE. 8 (6): e64752. Bibcode:2013PLoSO...864752H. doi: 10.1371/journal.pone.0064752 . ISSN 1932-6203. PMC 3673968 . PMID 23755141.
- ↑ Yamauchi, Takeshi; Yamasaki, Kenshi; Tsuchiyama, Kenichiro; Koike, Saaya; Aiba, Setsuya (June 2017). "A quantitative analysis of multilineage-differentiating stress-enduring (Muse) cells in human adipose tissue and efficacy of melanocytes induction". Journal of Dermatological Science. 86 (3): 198–205. doi: 10.1016/j.jdermsci.2017.03.001 . ISSN 1873-569X. PMID 28292562.
- ↑ Yamauchi, Takeshi; Yamasaki, Kenshi; Tsuchiyama, Kenichiro; Koike, Saaya; Aiba, Setsuya (2017-07-20). "The potential of Muse cells for regenerative medicine of skin: procedures to reconstitute skin with Muse cell-derived keratinocytes, fibroblasts, and melanocytes". The Journal of Investigative Dermatology. 137 (12): 2639–2642. doi: 10.1016/j.jid.2017.06.021 . ISSN 1523-1747. PMID 28736234.
- 1 2 Wakao, Shohei; Kitada, Masaaki; Kuroda, Yasumasa; Shigemoto, Taeko; Matsuse, Dai; Akashi, Hideo; Tanimura, Yukihiro; Tsuchiyama, Kenichiro; Kikuchi, Tomohiko (2011-06-14). "Multilineage-differentiating stress-enduring (Muse) cells are a primary source of induced pluripotent stem cells in human fibroblasts". Proceedings of the National Academy of Sciences of the United States of America. 108 (24): 9875–9880. Bibcode:2011PNAS..108.9875W. doi: 10.1073/pnas.1100816108 . ISSN 1091-6490. PMC 3116385 . PMID 21628574.
- 1 2 3 Iseki, Masahiro; Kushida, Yoshihiro; Wakao, Shohei; Akimoto, Takahiro; Mizuma, Masamichi; Motoi, Fuyuhiko; Asada, Ryuta; Shimizu, Shinobu; Unno, Michiaki (2017-05-09). "Muse Cells, Nontumorigenic Pluripotent-Like Stem Cells, Have Liver Regeneration Capacity Through Specific Homing and Cell Replacement in a Mouse Model of Liver Fibrosis". Cell Transplantation. 26 (5): 821–840. doi:10.3727/096368916X693662. ISSN 1555-3892. PMC 5657714 . PMID 27938474.
- 1 2 3 Katagiri, H.; Kushida, Y.; Nojima, M.; Kuroda, Y.; Wakao, S.; Ishida, K.; Endo, F.; Kume, K.; Takahara, T. (February 2016). "A Distinct Subpopulation of Bone Marrow Mesenchymal Stem Cells, Muse Cells, Directly Commit to the Replacement of Liver Components". American Journal of Transplantation. 16 (2): 468–483. doi: 10.1111/ajt.13537 . ISSN 1600-6143. PMID 26663569.
- ↑ Amin, Mohamed; Kushida, Yoshihiro; Wakao, Shohei; Kitada, Masaaki; Tatsumi, Kazuki; Dezawa, Mari (2018). "Cardiotrophic Growth Factor–Driven Induction of Human Muse Cells into Cardiomyocyte-Like Phenotype". Cell Transplantation. 27 (2): 285–298. doi:10.1177/0963689717721514. PMC 5898685 . PMID 29637816.
- ↑ Yamada, Yoshihisa; Minatoguchi, Shingo; Baba, Shinya; Shibata, Sanae; Takashima, Satoshi; Wakao, Shohei; Okura, Hiroyuki; Dezawa, Mari; Minatoguchi, Shinya (2022). "Human Muse cells reduce myocardial infarct size and improve cardiac function without causing arrythmias in a swine model of acute myocardial infarction". PLOS ONE. 17 (3): e0265347. Bibcode:2022PLoSO..1765347Y. doi: 10.1371/journal.pone.0265347 . ISSN 1932-6203. PMC 8947423 . PMID 35324926.
- 1 2 Uchida, Hiroki; Niizuma, Kuniyasu; Kushida, Yoshihiro; Wakao, Shohei; Tominaga, Teiji; Borlongan, Cesario V; Dezawa, Mari (2017). "Human Muse Cells Reconstruct Neuronal Circuitry in Subacute Lacunar Stroke Model". Stroke. 48 (2): 428–435. doi:10.1161/STROKEAHA.116.014950. PMC 5262965 . PMID 27999136.
- ↑ Shimamura, Norihito; Kakuta, Kiyohide; Wang, Liang; Naraoka, Masato; Uchida, Hiroki; Wakao, Shohei; Dezawa, Mari; Ohkuma, Hiroki (February 2017). "Neuro-regeneration therapy using human Muse cells is highly effective in a mouse intracerebral hemorrhage model". Experimental Brain Research. 235 (2): 565–572. doi:10.1007/s00221-016-4818-y. ISSN 1432-1106. PMID 27817105. S2CID 11415086.
- ↑ Shono, Yoshihiro; Kushida, Yoshihiro; Wakao, Shohei; Kuroda, Yasumasa; Unno, Michiaki; Kamei, Takashi; Miyagi, Shigehito; Dezawa, Mari (June 2021). "Protection of liver sinusoids by intravenous administration of human Muse cells in a rat extra-small partial liver transplantation model". American Journal of Transplantation: Official Journal of the American Society of Transplantation and the American Society of Transplant Surgeons. 21 (6): 2025–2039. doi:10.1111/ajt.16461. ISSN 1600-6143. PMC 8248424 . PMID 33350582.
- ↑ Kinoshita, Kahori; Kuno, Shinichiro; Ishimine, Hisako; Aoi, Noriyuki; Mineda, Kazuhide; Kato, Harunosuke; Doi, Kentaro; Kanayama, Koji; Feng, Jingwei (February 2015). "Therapeutic Potential of Adipose-Derived SSEA-3-Positive Muse Cells for Treating Diabetic Skin Ulcers". Stem Cells Translational Medicine. 4 (2): 146–155. doi:10.5966/sctm.2014-0181. ISSN 2157-6564. PMC 4303359 . PMID 25561682.
- ↑ Hosoyama, Katsuhiro; Wakao, Shohei; Kushida, Yoshihiro; Ogura, Fumitaka; Maeda, Kay; Adachi, Osamu; Kawamoto, Shunsuke; Dezawa, Mari; Saiki, Yoshikatsu (2018). "Intravenously injected human multilineage-differentiating stress-enduring cells selectively engraft into mouse aortic aneurysms and attenuate dilatation by differentiating into multiple cell types". The Journal of Thoracic and Cardiovascular Surgery. 155 (6): 2301–2313.e4. doi: 10.1016/j.jtcvs.2018.01.098 . PMID 29559260.
- ↑ Suzuki, Toshihiko; Sato, Yoshiaki; Kushida, Yoshihiro; Tsuji, Masahiro; Wakao, Shohei; Ueda, Kazuto; Imai, Kenji; Iitani, Yukako; Shimizu, Shinobu; Hida, Hideki; Temma, Takashi; Saito, Shigeyoshi; Iida, Hidehiro; Mizuno, Masaaki; Takahashi, Yoshiyuki; Dezawa, Mari; Borlongan, Cesar V; Hayakawa, Masahiro (2020-11-22). "Intravenously delivered multilineage-differentiating stress enduring cells dampen excessive glutamate metabolism and microglial activation in experimental perinatal hypoxic ischemic encephalopathy". Journal of Cerebral Blood Flow & Metabolism. 41 (7): 1707–1720. doi: 10.1177/0271678X20972656 . PMC 8217885 . PMID 33222596. S2CID 227133496.
- ↑ Yamashita, Toru; Kushida, Yoshihiro; Wakao, Shohei; Tadokoro, Koh; Nomura, Emi; Omote, Yoshio; Takemoto, Mami; Hishikawa, Nozomi; Ohta, Yasuyuki; Dezawa, Mari; Abe, Koji (2020-10-13). "Therapeutic benefit of Muse cells in a mouse model of amyotrophic lateral sclerosis" (PDF). Scientific Reports. 10 (1): 17102. doi:10.1038/s41598-020-74216-4. PMC 554047 . PMID 33051552. S2CID 222344158.
- ↑ Ozuru, Ryo; Wakao, Shohei; Tsuji, Takahiro; Ohara, Naoya; Matsuba, Takashi; Amuran, Muhammad Y; Isobe, Junko; Iino, Morio; Nishida, Naoki; Matsumoto, Sari; Iwadate, Kimiharu; Konishi, Noriko; Yasuda, Kaori; Tashiro, Kosuke; Hida, Misato; Yadoiwa, Arisato; Kato, Shinsuke; Yamashita, Eijiro; Matsumoto, Sohkichi; Kurozawa, Yoichi; Dezawa, Mari; Fujii, Jun (2019-10-01). "Rescue from Stx2-Producing E. coli-Associated Encephalopathy by Intravenous Injection of Muse Cells in NOD-SCID Mice" (PDF). American Society of Gene & Cell Therapy. 28 (1): 100–118. doi: 10.1016/j.ymthe.2019.09.023 . PMC 6953779 . PMID 31607541. S2CID 204544398.
- ↑ Dushime, Honorine; Moreno, Stéphanie G.; Linard, Christine; Adrait, Annie; Couté, Yohann; Peltzer, Juliette; Messiaen, Sébastien; Torres, Claire; Bensemmane, Lydia; Lewandowski, Daniel; Romeo, Paul-Henri; Petit, Vanessa; Gault, Nathalie (2023-08-11). "Fetal Muse-based therapy prevents lethal radio-induced gastrointestinal syndrome by intestinal regeneration". Stem Cell Research & Therapy. 14 (1): 201. doi: 10.1186/s13287-023-03425-1 . ISSN 1757-6512. PMC 10416451 . PMID 37568164.
- 1 2 Hori, Emiko; Hayakawa, Yumiko; Hayashi, Tomohide; Hori, Satoshi; Okamoto, Soushi; Shibata, Takashi; Kubo, Michiya; Horie, Yukio; Sasahara, Masakiyo (June 2016). "Mobilization of Pluripotent Multilineage-Differentiating Stress-Enduring Cells in Ischemic Stroke". Journal of Stroke and Cerebrovascular Diseases. 25 (6): 1473–1481. doi:10.1016/j.jstrokecerebrovasdis.2015.12.033. ISSN 1532-8511. PMID 27019988.
- ↑ Tanaka, Toshiki; Nishigaki, Kazuhiko; Minatoguchi, Shingo; Nawa, Takahide; Yamada, Yoshihisa; Kanamori, Hiromitsu; Mikami, Atsushi; Ushikoshi, Hiroaki; Kawasaki, Masanori; Dezawa, Mari; Minatoguchi, Shinya (2018). "Mobilized Muse Cells After Acute Myocardial Infarction Predict Cardiac Function and Remodeling in the Chronic Phase". Circulation Journal. 82 (2): 561–571. doi: 10.1253/circj.CJ-17-0552 . hdl: 20.500.12099/74896 . PMID 28931784.
- ↑ Cosset, Jean Marc (2002). "ESTRO Breur Gold Medal Award Lecture 2001: Irradiation accidents – lessons for oncology?". Radiotherapy and Oncology. 63 (1): 1–10. doi:10.1016/s0167-8140(02)00059-2. PMID 12065098.
- ↑ Kuroda, Yasumasa; Kitada, Masaaki; Wakao, Shohei; Dezawa, Mari (2011). "Bone Marrow Mesenchymal Cells: How Do They Contribute to Tissue Repair and Are They Really Stem Cells?". Archivum Immunologiae et Therapiae Experimentalis. 59 (5): 369–78. doi:10.1007/s00005-011-0139-9. PMID 21789625. S2CID 12073078.
- ↑ "Life Science Institute, Inc. initiates clinical trials of Muse-cell product CL2020 to treat acute respiratory distress syndrome (ARDS) related to novel coronavirus (SARS-CoV-2) infection" (PDF). LSII. 2021-04-20. Archived from the original (PDF) on 2021-04-23. Retrieved 2024-03-19.
- ↑ Matsuyama, Nao; Shimizu, Shinobu; Ueda, Kazuto; Suzuki, Toshihiko; Suzuki, Sakiko; Miura, Ryosuke; Katayama, Akemi; Ando, Masahiko; Mizuno, Masaaki; Hirakawa, Akihiro; Hayakawa, Masahiro; Sato, Yoshiaki (2022-04-26). "Safety and tolerability of a multilineage-differentiating stress-enduring cell-based product in neonatal hypoxic-ischaemic encephalopathy with therapeutic hypothermia (SHIELD trial): a clinical trial protocol open-label, non-randomised, dose-escalation trial". BMJ Open. 12 (4): e057073. doi:10.1136/bmjopen-2021-057073. ISSN 2044-6055. PMC 9045108 . PMID 35473726.