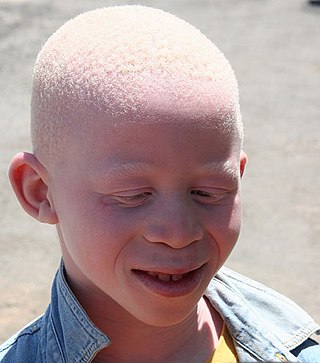
Albinism is a congenital condition characterized in humans by the partial or complete absence of pigment in the skin, hair and eyes. Albinism is associated with a number of vision defects, such as photophobia, nystagmus, and amblyopia. Lack of skin pigmentation makes for more susceptibility to sunburn and skin cancers. In rare cases such as Chédiak–Higashi syndrome, albinism may be associated with deficiencies in the transportation of melanin granules. This also affects essential granules present in immune cells, leading to increased susceptibility to infection.
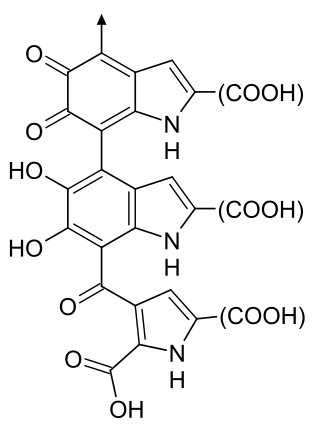
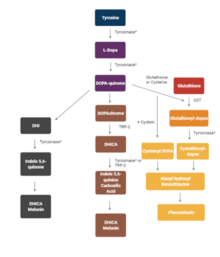
Melanin consist of oligomers or polymers arranged in a disordered manner which among other functions provide the pigments of many organisms. Melanin pigments are produced in a specialized group of cells known as melanocytes. They have been described as "among the last remaining biological frontiers with the unknown".

Melanocytes are melanin-producing neural crest-derived cells located in the bottom layer of the skin's epidermis, the middle layer of the eye, the inner ear, vaginal epithelium, meninges, bones, and heart. Melanin is a dark pigment primarily responsible for skin color. Once synthesized, melanin is contained in special organelles called melanosomes which can be transported to nearby keratinocytes to induce pigmentation. Thus darker skin tones have more melanosomes present than lighter skin tones. Functionally, melanin serves as protection against UV radiation. Melanocytes also have a role in the immune system.

A melanosome is an organelle found in animal cells and is the site for synthesis, storage and transport of melanin, the most common light-absorbing pigment found in the animal kingdom. Melanosomes are responsible for color and photoprotection in animal cells and tissues.

Browning is the process of food turning brown due to the chemical reactions that take place within. The process of browning is one of the chemical reactions that take place in food chemistry and represents an interesting research topic regarding health, nutrition, and food technology. Though there are many different ways food chemically changes over time, browning in particular falls into two main categories: enzymatic versus non-enzymatic browning processes.

Hypopigmentation is characterized specifically as an area of skin becoming lighter than the baseline skin color, but not completely devoid of pigment. This is not to be confused with depigmentation, which is characterized as the absence of all pigment. It is caused by melanocyte or melanin depletion, or a decrease in the amino acid tyrosine, which is used by melanocytes to make melanin. Some common genetic causes include mutations in the tyrosinase gene or OCA2 gene. As melanin pigments tend to be in the skin, eye, and hair, these are the commonly affected areas in those with hypopigmentation.

Skin whitening, also known as skin lightening and skin bleaching, is the practice of using chemical substances in an attempt to lighten the skin or provide an even skin color by reducing the melanin concentration in the skin. Several chemicals have been shown to be effective in skin whitening, while some have proven to be toxic or have questionable safety profiles. This includes mercury compounds which may cause neurological problems and kidney problems.
Catechol oxidase is a copper oxidase that contains a type 3 di-copper cofactor and catalyzes the oxidation of ortho-diphenols into ortho-quinones coupled with the reduction of molecular oxygen to water. It is present in a variety of species of plants and fungi including Ipomoea batatas and Camellia sinensis. Metalloenzymes with type 3 copper centers are characterized by their ability to reversibly bind dioxygen at ambient conditions. In plants, catechol oxidase plays a key role in enzymatic browning by catalyzing the oxidation of catechol to o-quinone in the presence of oxygen, which can rapidly polymerize to form the melanin that grants damaged fruits their dark brown coloration.

Agouti-signaling protein is a protein that in humans is encoded by the ASIP gene. It is responsible for the distribution of melanin pigment in mammals. Agouti interacts with the melanocortin 1 receptor to determine whether the melanocyte produces phaeomelanin, or eumelanin. This interaction is responsible for making distinct light and dark bands in the hairs of animals such as the agouti, which the gene is named after. In other species such as horses, agouti signalling is responsible for determining which parts of the body will be red or black. Mice with wildtype agouti will be grey-brown, with each hair being partly yellow and partly black. Loss of function mutations in mice and other species cause black fur coloration, while mutations causing expression throughout the whole body in mice cause yellow fur and obesity.

A white horse is born predominantly white and stays white throughout its life. A white horse has mostly pink skin under its hair coat, and may have brown, blue, or hazel eyes. "True white" horses, especially those that carry one of the dominant white (W) genes, are rare. Most horses that are commonly referred to as "white" are actually "gray" horses whose hair coats are completely white. Gray horses may be born of any color and their hairs gradually turn white as time goes by and take on a white appearance. Nearly all gray horses have dark skin, except under any white markings present at birth. Skin color is the most common method for an observer to distinguish between mature white and gray horses.

Microphthalmia-associated transcription factor also known as class E basic helix-loop-helix protein 32 or bHLHe32 is a protein that in humans is encoded by the MITF gene.
Polyphenol oxidase, an enzyme involved in fruit browning, is a tetramer that contains four atoms of copper per molecule.

The genetic basis of coat colour in the Labrador Retriever has been found to depend on several distinct genes. The interplay among these genes is used as an example of epistasis.

Tyrosinase-related protein 1, also known as TYRP1, is an intermembrane enzyme which in humans is encoded by the TYRP1 gene.

P protein, also known as melanocyte-specific transporter protein or pink-eyed dilution protein homolog, is a protein that in humans is encoded by the oculocutaneous albinism II (OCA2) gene. The P protein is believed to be an integral membrane protein involved in small molecule transport, specifically of tyrosine—a precursor of melanin. Certain mutations in OCA2 result in type 2 oculocutaneous albinism. OCA2 encodes the human homologue of the mouse p gene.

Membrane-associated transporter protein (MATP), also known as solute carrier family 45 member 2 (SLC45A2) or melanoma antigen AIM1, is a protein that in humans is encoded by the SLC45A2 gene.
Oculocutaneous albinism type I or type 1A is an autosomal recessive skin disease. This subtype of oculocutaneous albinism is caused when the gene for tyrosinase does not function properly.

Ocular albinism type 1(OA1) is the most common type of ocular albinism, with a prevalence rate of 1:50,000. It is an inheritable classical Mendelian type X-linked recessive disorder wherein the retinal pigment epithelium lacks pigment while hair and skin appear normal. Since it is usually an X-linked disorder, it occurs mostly in males, while females are carriers unless they are homozygous. About 60 missense and nonsense mutations, insertions, and deletions have been identified in Oa1. Mutations in OA1 have been linked to defective glycosylation and thus improper intracellular transportation.

Amelanism is a pigmentation abnormality characterized by the lack of pigments called melanins, commonly associated with a genetic loss of tyrosinase function. Amelanism can affect fish, amphibians, reptiles, birds, and mammals including humans. The appearance of an amelanistic animal depends on the remaining non-melanin pigments. The opposite of amelanism is melanism, a higher percentage of melanin.

Albinism is the congenital absence of melanin in an animal or plant resulting in white hair, feathers, scales and skin and reddish pink or blue eyes. Individuals with the condition are referred to as albinos.