Related Research Articles

The olfactory bulb is a neural structure of the vertebrate forebrain involved in olfaction, the sense of smell. It sends olfactory information to be further processed in the amygdala, the orbitofrontal cortex (OFC) and the hippocampus where it plays a role in emotion, memory and learning. The bulb is divided into two distinct structures: the main olfactory bulb and the accessory olfactory bulb. The main olfactory bulb connects to the amygdala via the piriform cortex of the primary olfactory cortex and directly projects from the main olfactory bulb to specific amygdala areas. The accessory olfactory bulb resides on the dorsal-posterior region of the main olfactory bulb and forms a parallel pathway. Destruction of the olfactory bulb results in ipsilateral anosmia, while irritative lesions of the uncus can result in olfactory and gustatory hallucinations.
Stimulus modality, also called sensory modality, is one aspect of a stimulus or what is perceived after a stimulus. For example, the temperature modality is registered after heat or cold stimulate a receptor. Some sensory modalities include: light, sound, temperature, taste, pressure, and smell. The type and location of the sensory receptor activated by the stimulus plays the primary role in coding the sensation. All sensory modalities work together to heighten stimuli sensation when necessary.

The olfactory system, or sense of smell, is the sensory system used for smelling (olfaction). Olfaction is one of the special senses, that have directly associated specific organs. Most mammals and reptiles have a main olfactory system and an accessory olfactory system. The main olfactory system detects airborne substances, while the accessory system senses fluid-phase stimuli.

The olfactory epithelium is a specialized epithelial tissue inside the nasal cavity that is involved in smell. In humans, it measures 5 cm2 (0.78 sq in) and lies on the roof of the nasal cavity about 7 cm (2.8 in) above and behind the nostrils. The olfactory epithelium is the part of the olfactory system directly responsible for detecting odors.

Olfactory receptors (ORs), also known as odorant receptors, are chemoreceptors expressed in the cell membranes of olfactory receptor neurons and are responsible for the detection of odorants which give rise to the sense of smell. Activated olfactory receptors trigger nerve impulses which transmit information about odor to the brain. These receptors are members of the class A rhodopsin-like family of G protein-coupled receptors (GPCRs). The olfactory receptors form a multigene family consisting of around 800 genes in humans and 1400 genes in mice.
Body odor or body odour (BO) is present in all animals and its intensity can be influenced by many factors. Body odor has a strong genetic basis, but can also be strongly influenced by various factors, such as gender, diet, health, and medication. The body odor of human males plays an important role in human sexual attraction, as a powerful indicator of MHC/HLA heterozygosity. Significant evidence suggests that women are attracted to men whose body odor is different from theirs, indicating that they have immune genes that are different from their own, which may produce healthier offspring.
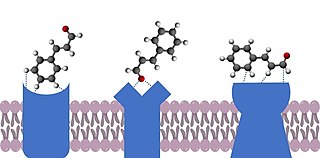
The docking theory of olfaction proposes that the smell of an odorant molecule is due to a range of weak non-covalent interactions between the odorant [a ligand] and one or more G protein-coupled odorant receptors. These include intermolecular forces, such as dipole-dipole and Van der Waals interactions, as well as hydrogen bonding. More specific proposed interactions include metal-ion, ion-ion, cation-pi and pi-stacking. Interactions can be influenced by the hydrophobic effect. Conformational changes can also have a significant impact on interactions with receptors, as ligands have been shown to interact with ligands without being in their conformation of lowest energy.
The vibration theory of smell proposes that a molecule's smell character is due to its vibrational frequency in the infrared range. This controversial theory is an alternative to the more widely accepted docking theory of olfaction, which proposes that a molecule's smell character is due to a range of weak non-covalent interactions between its protein odorant receptor, such as electrostatic and Van der Waals interactions as well as H-bonding, dipole attraction, pi-stacking, metal ion, Cation–pi interaction, and hydrophobic effects, in addition to the molecule's conformation.
A sensory cue is a statistic or signal that can be extracted from the sensory input by a perceiver, that indicates the state of some property of the world that the perceiver is interested in perceiving.
Olfactory fatigue, also known as odor fatigue, olfactory adaptation, and noseblindness, is the temporary, normal inability to distinguish a particular odor after a prolonged exposure to that airborne compound. For example, when entering a restaurant initially the odor of food is often perceived as being very strong, but after time the awareness of the odor normally fades to the point where the smell is not perceptible or is much weaker. After leaving the area of high odor, the sensitivity is restored with time. Anosmia is the permanent loss of the sense of smell, and is different from olfactory fatigue.
Dysosmia is a disorder described as any qualitative alteration or distortion of the perception of smell. Qualitative alterations differ from quantitative alterations, which include anosmia and hyposmia. Dysosmia can be classified as either parosmia or phantosmia. Parosmia is a distortion in the perception of an odorant. Odorants smell different from what one remembers. Phantosmia is the perception of an odor when no odorant is present. The cause of dysosmia still remains a theory. It is typically considered a neurological disorder and clinical associations with the disorder have been made. Most cases are described as idiopathic and the main antecedents related to parosmia are URTIs, head trauma, and nasal and paranasal sinus disease. Dysosmia tends to go away on its own but there are options for treatment for patients that want immediate relief.

An odor or odour is caused by one or more volatilized chemical compounds that are generally found in low concentrations that humans and many animals can perceive via their sense of smell. An odor is also called a "smell" or a "scent", which can refer to either a pleasant or an unpleasant odor.

The sense of smell, or olfaction, is the special sense through which smells are perceived. The sense of smell has many functions, including detecting desirable foods, hazards, and pheromones, and plays a role in taste.
Imprinting is most often used to describe an early-life bond that can later affect an animal's mate choice. More broadly, the term refers to a rapid and selective learning process that only can occur during certain times in an animal's life. In sheep, ewes having just given birth imprint onto their lambs on the basis of olfactory cues, allowing mothers to distinguish their own offspring from other lambs in the flock. This olfactory-based imprinting is dependent on a ewe's behavior after giving birth, on the presence of amniotic fluid, and on a specialized odor-influenced learning process which allows the ewe to quickly memorize the smell of her offspring, to whom she then forms an exclusive maternal bond.

Sniffing is a perceptually-relevant behavior, defined as the active sampling of odors through the nasal cavity for the purpose of information acquisition. This behavior, displayed by all terrestrial vertebrates, is typically identified based upon changes in respiratory frequency and/or amplitude, and is often studied in the context of odor guided behaviors and olfactory perceptual tasks. Sniffing is quantified by measuring intra-nasal pressure or flow or air or, while less accurate, through a strain gauge on the chest to measure total respiratory volume. Strategies for sniffing behavior vary depending upon the animal, with small animals displaying sniffing frequencies ranging from 4 to 12 Hz but larger animals (humans) sniffing at much lower frequencies, usually less than 2 Hz. Subserving sniffing behaviors, evidence for an "olfactomotor" circuit in the brain exists, wherein perception or expectation of an odor can trigger brain respiratory center to allow for the modulation of sniffing frequency and amplitude and thus acquisition of odor information. Sniffing is analogous to other stimulus sampling behaviors, including visual saccades, active touch, and whisker movements in small animals. Atypical sniffing has been reported in cases of neurological disorders, especially those disorders characterized by impaired motor function and olfactory perception.
Odor molecules are detected by the olfactory receptors in the olfactory epithelium of the nasal cavity. Each receptor type is expressed within a subset of neurons, from which they directly connect to the olfactory bulb in the brain. Olfaction is essential for survival in most vertebrates; however, the degree to which an animal depends on smell is highly varied. Great variation exists in the number of OR genes among vertebrate species, as shown through bioinformatic analyses. This diversity exists by virtue of the wide-ranging environments that they inhabit. For instance, dolphins that are secondarily adapted to an aquatic niche possess a considerably smaller subset of genes than most mammals. OR gene repertoires have also evolved in relation to other senses, as higher primates with well-developed vision systems tend to have a smaller number of OR genes. As such, investigating the evolutionary changes of OR genes can provide useful information on how genomes respond to environmental changes. Differences in smell sensitivity are also dependent on the anatomy of the olfactory apparatus, such as the size of the olfactory bulb and epithelium.

Insect olfaction refers to the function of chemical receptors that enable insects to detect and identify volatile compounds for foraging, predator avoidance, finding mating partners and locating oviposition habitats. Thus, it is the most important sensation for insects. Most important insect behaviors must be timed perfectly which is dependent on what they smell and when they smell it. For example, olfaction is essential for locating host plants and hunting prey in many species of insects, such as the moth Deilephila elpenor and the wasp Polybia sericea, respectively.
Retronasal smell, retronasal olfaction, is the ability to perceive flavor dimensions of foods and drinks. Retronasal smell is a sensory modality that produces flavor. It is best described as a combination of traditional smell and taste modalities. Retronasal smell creates flavor from smell molecules in foods or drinks shunting up through the nasal passages as one is chewing. When people use the term "smell", they are usually referring to "orthonasal smell", or the perception of smell molecules that enter directly through the nose and up the nasal passages. Retronasal smell is critical for experiencing the flavor of foods and drinks. Flavor should be contrasted with taste, which refers to five specific dimensions: (1) sweet, (2) salty, (3) bitter, (4) sour, and (5) umami. Perceiving anything beyond these five dimensions, such as distinguishing the flavor of an apple from a pear for example, requires the sense of retronasal smell.
Neurogastronomy is the study of flavor perception and the ways it affects cognition and memory. This interdisciplinary field is influenced by the psychology and neuroscience of sensation, learning, satiety, and decision making. Areas of interest include how olfaction contributes to flavor, food addiction and obesity, taste preferences, and the linguistics of communicating and identifying flavor. The term neurogastronomy was coined by neuroscientist Gordon M. Shepherd.

Olfactic communication is a channel of nonverbal communication referring to the various ways people and animals communicate and engage in social interaction through their sense of smell. Our human olfactory sense is one of the most phylogenetically primitive and emotionally intimate of the five senses; the sensation of smell is thought to be the most matured and developed human sense.
References
- 1 2 3 4 Wilson, DA (2003). "The fundamental role of memory in olfactory perception". Trends in Neurosciences. 26 (5): 243–7. CiteSeerX 10.1.1.314.8907 . doi:10.1016/s0166-2236(03)00076-6. PMID 12744840. S2CID 10433103.
- ↑ Pinel, J.P. (2006). Biopsychology. 6th ed. Boston, MA, US: Pearson Education Inc.
- ↑ Guerin, D. (2008). Norepinephrine neuromodulation in the olfactory bulb modulates odor habituation and spontaneous discrimination. Behavioral neuroscience, 122(4), 816.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Wilson, DA. (2006). Learning to smell: Olfactory perception from neurobiology to behavior.. Baltimore, MD, US: Johns Hopkins University Press.
- ↑ Guerin, D. (2008). Noradrenergic neuromodulation in the olfactory bulb modulates odor habituation and spontaneous discrimination. Behavioral neuroscience, 122(4), 824.
- ↑ De Rosa, Eve. (2000). Muscarinic Cholinergic Neuromodulation Reduces Proactive Interference Between Stored Odor Memories During Associative Learning in Rats. Beahvioural Neuroscience, 114(1), 29-40.
- 1 2 3 4 Rouby, C., Schaal, B., Dubois, D., Gervais, R., & Holley, A., (Eds.). (2002). Olfaction, taste and cognition. New York: Cambridge University Press.
- 1 2 3 4 5 6 7 Schab, F., & Crowder, R. G. (Eds.). (1995). Memory for odors. Mahwah, NJ: Lawrence Erlbaum Associates, Inc.
- 1 2 3 4 5 Schab, FR (1991). "Odor memory: Taking stock". Psychological Bulletin. 109 (2): 242–251. doi:10.1037/0033-2909.109.2.242. PMID 2034752.
- ↑ Radvansky, G., (2006). Human Memory. Boston, MA: Pearson Education Group, Inc.
- 1 2 3 4 5 Olsson, MJ. (2003). Implicit and explicit memory for odors: Hemispheric differences. Memory & cognition, 31(1), 44-50.
- ↑ Hummel, Thomas; Rissom, Karo; Reden, Jens; Hähner, Aantje; Weidenbecher, Mark; Hüttenbrink, Karl-Bernd (March 2009). "Effects of olfactory training in patients with olfactory loss". The Laryngoscope. 119 (3): 496–499. doi:10.1002/lary.20101. ISSN 1531-4995. PMID 19235739. S2CID 5239574.
- 1 2 3 4 5 Purves, D. Augustine, G. Fitzpatrick, D. Hall, W. LaMantia, A.S. White, L. (2012). Neuroscience 5th Edition. Sunderland, MA: Sinauer
- ↑ Buchanan, TW. (2003). A specific role for the human amygdala in olfactory memory. Learning & memory, 10(5), P 319.
- ↑ Herz RS, Engen T (September 1996). "Odor memory: Review and analysis". Psychonomic Bulletin & Review. 3 (3): 300–313. doi: 10.3758/BF03210754 . PMID 24213931.
- ↑ Herz, Rachel S. (2016). "The Role of Odor-Evoked Memory in Psychological and Physiological Health". Brain Sciences. 6 (3): 22. doi: 10.3390/brainsci6030022 . PMC 5039451 . PMID 27447673.
 Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License. - ↑ Herz, R.S., Eliassen, J., Beland, S. and Souza, T. (2004). evidence for the emotional potency of odor-evoked memory, 42(3), pp.371–378.
- 1 2 3 Lévy, F., Locatelli, A., Piketty, V., Tillet, Y., & Poindron, P. (1994). Involvement of the main but not the accessory olfactory system in maternal behavior of primiparous and multiparous ewes. Physiology and Behavior (57) 1: 97-104.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 Lévy, F.; Keller, M.; Poindron, P. (2003). "Olfactory regulation of maternal behavior in mammals". Hormones and Behavior. 46 (3): 284–302. doi:10.1016/j.yhbeh.2004.02.005. PMID 15325229. S2CID 31209629.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 Porter, R.H.; Winberg, J. (1997). "Unique salience of maternal breast odors for newborn infants". Neuroscience and Biobehavioral Reviews. 23 (3): 439–449. doi:10.1016/s0149-7634(98)00044-x. PMID 9989430. S2CID 34940555.
- 1 2 3 4 Broad, K.D., Hinton, M.R., Keverne, B., & Kendrick, K.M. (2002). Involvement of the medial prefrontal cortex in mediating behavioral responses to odor cues rather than olfactory recognition memory. Neuroscience (114) 5: 715-729.
- 1 2 3 4 5 6 7 8 9 Varendi, H.; Christensson, K.; Porter, H.; Winberg, J. (1997). "Soothing effect of amniotic fluid smell in newborn infants". Early Human Development. 51 (1): 47–55. doi:10.1016/s0378-3782(97)00082-0. PMID 9570031.
- ↑ Magill, Frank Northern. 1998. Psychology Basics. Pasadena, CA: Salem Press. P 418-419.
- ↑ Gazzaniga, Michael S. 1998. The Mind's Past. Berkeley, CA: University of California Press. P 105.
- 1 2 Goldstein, Bruce E. 2002. Sensation and Perception: 6th Edition. Pacific Grove CA: Wadsworth Group. P 477.
- 1 2 Goldstein, Bruce E. 2002. Sensation and Perception: 6th Edition. Pacific Grove CA: Wadsworth Group. P 475.
- 1 2 3 4 5 Stockhorst, Ursula; Pietrowsky, Reinhard (2004). "Olfactory perception, communication, and the nose-to-brain pathway". Physiology & Behavior. 83 (1): 3–11. doi:10.1016/j.physbeh.2004.07.018. PMID 15501485. S2CID 54433584.
- ↑ Kovács, Tibor (2004-04-01). "Mechanisms of olfactory dysfunction in aging and neurodegenerative disorders". Ageing Research Reviews. 3 (2): 215–232. doi:10.1016/j.arr.2003.10.003. PMID 15177056. S2CID 23252066.
- ↑ Cohen, Alex S.; Brown, Laura A.; Auster, Tracey L. (2012-03-01). "Olfaction, "olfiction," and the schizophrenia-spectrum: An updated meta-analysis on identification and acuity". Schizophrenia Research. 135 (1–3): 152–157. doi:10.1016/j.schres.2011.12.005. ISSN 0920-9964. PMID 22244185. S2CID 17159465.
- ↑ Brewer, Warrick J.; Wood, Stephen J.; McGorry, Patrick D.; Francey, Shona M.; Phillips, Lisa J.; Yung, Alison R.; Anderson, Vicki; Copolov, David L.; Singh, Bruce (2003-10-01). "Impairment of Olfactory Identification Ability in Individuals at Ultra-High Risk for Psychosis Who Later Develop Schizophrenia". American Journal of Psychiatry. 160 (10): 1790–1794. doi:10.1176/appi.ajp.160.10.1790. ISSN 0002-953X. PMID 14514492. S2CID 26789096.
- ↑ Moberg, Paul J.; Agrin, Rachel; Gur, Raquel E.; Gur, Ruben C.; Turetsky, Bruce I.; Doty, Richard L. (September 1999). "Olfactory Dysfunction in Schizophrenia: A Qualitative and Quantitative Review". Neuropsychopharmacology. 21 (3): 325–340. doi: 10.1016/S0893-133X(99)00019-6 . ISSN 0893-133X. PMID 10457530.
- ↑ Doty, Richard (2012). "Olfaction in Parkinson's disease and related disorders". Neurobiological Disorders. 46 (3): 527–552. doi:10.1016/j.nbd.2011.10.026. PMC 3429117 . PMID 22192366.
- ↑ Wattendorf, Elise; Welge-Lüssen, Antje; Fiedler, Klaus; Bilecen, Deniz; Wolfensberger, Markus; Fuhr, Peter; Hummel, Thomas; Westermann, Birgit (2009-12-09). "Olfactory Impairment Predicts Brain Atrophy in Parkinson9s Disease". Journal of Neuroscience. 29 (49): 15410–15413. doi:10.1523/JNEUROSCI.1909-09.2009. ISSN 0270-6474. PMC 6666111 . PMID 20007465.
- 1 2 Mesholam, Raquelle I.; Moberg, Paul J.; Mahr, Richard N.; Doty, Richard L. (1998-01-01). "Olfaction in Neurodegenerative Disease: A Meta-analysis of Olfactory Functioning in Alzheimer's and Parkinson's Diseases". Archives of Neurology. 55 (1): 84–90. doi:10.1001/archneur.55.1.84. ISSN 0003-9942. PMID 9443714.
- ↑ Moberg, Paul J.; Pearlson, Godfrey D.; Speedie, Lynn J.; Lipsey, John R.; Strauss, Milton E.; Folstein, Susan E. (1987-12-01). "Olfactory Recognition: Differential Impairments in Early and Late Huntington's and Alzheimer's Diseases". Journal of Clinical and Experimental Neuropsychology. 9 (6): 650–664. doi:10.1080/01688638708405208. ISSN 0168-8634. PMID 2961789.
- ↑ Nordin, Steven; Paulsen, Jane S.; Murphy, Claire (May 1995). "Sensory- and memory-mediated olfactory dysfunction in Huntington's disease". Journal of the International Neuropsychological Society. 1 (3): 281–290. doi:10.1017/S1355617700000278. ISSN 1469-7661. PMID 9375222. S2CID 32948555.
- ↑ Mair, R. G.; Doty, R. L.; Kelly, K. M.; Wilson, C. S.; Langlais, P. J.; McEntee, W. J.; Vollmecke, T. A. (1986-01-01). "Multimodal sensory discrimination deficits in Korsakoff's psychosis". Neuropsychologia. 24 (6): 831–839. doi:10.1016/0028-3932(86)90082-5. PMID 2433640. S2CID 17814461.
- ↑ Jones, Barbara P.; Moskowitz, Howard R.; Butters, Nelson (1975-04-01). "Olfactory discrimination in alcoholic korsakoff patients". Neuropsychologia. 13 (2): 173–179. doi:10.1016/0028-3932(75)90026-3. PMID 1153100. S2CID 31712440.
- ↑ Doty, Richard L.; Reyes, Patricio F.; Gregor, Tom (1987-05-01). "Presence of both odor identification and detection deficits in alzheimer's disease". Brain Research Bulletin. 18 (5): 597–600. doi:10.1016/0361-9230(87)90129-8. PMID 3607528. S2CID 4766725.
- ↑ Devanand, D. P.; Lee, Seonjoo; Manly, Jennifer; Andrews, Howard; Schupf, Nicole; Doty, Richard L.; Stern, Yaakov; Zahodne, Laura B.; Louis, Elan D. (2015-01-13). "Olfactory deficits predict cognitive decline and Alzheimer dementia in an urban community". Neurology. 84 (2): 182–189. doi:10.1212/wnl.0000000000001132. ISSN 0028-3878. PMC 4336090 . PMID 25471394.
- ↑ Wilson, Robert S.; Arnold, Steven E.; Schneider, Julie A.; Boyle, Patricia A.; Buchman, Aron S.; Bennett, David A. (2009-07-01). "Olfactory Impairment in Presymptomatic Alzheimer's Disease". Annals of the New York Academy of Sciences. 1170 (1): 730–735. Bibcode:2009NYASA1170..730W. doi:10.1111/j.1749-6632.2009.04013.x. ISSN 1749-6632. PMC 2857767 . PMID 19686220.
- ↑ Attems, J.; Jellinger, K. A. (November 2006). "Olfactory tau pathology in Alzheimer disease and mild cognitive impairment". Clinical Neuropathology. 25 (6): 265–271. ISSN 0722-5091. PMID 17140156.
- 1 2 3 Lamboin. S et al. 2007. Effects of anti-depressents on olfactory sensitivity in mice. Progress in Neuro-Psychopharmacology & Biological Psychiatry 32 (2008) 629–632.
- ↑ Doy, Richard L.; Newhouse, Marisa G.; Azzalina, Jeffrey D. (1985-01-01). "Internal consistency and short-term test-retest reliability of the University of Pennsylvania Smell Identification Test". Chemical Senses. 10 (3): 297–300. doi:10.1093/chemse/10.3.297. ISSN 0379-864X.
- ↑ Doty, Richard L.; Marcus, Avron; William Lee, W. (1996-03-01). "Development of the 12-Item Cross-Cultural Smell Identification Test(CC-SIT)". The Laryngoscope. 106 (3): 353–356. doi:10.1097/00005537-199603000-00021. ISSN 1531-4995. PMID 8614203. S2CID 24605655.
- ↑ Hummel, T.; Sekinger, B.; Wolf, S. R.; Pauli, E.; Kobal, G. (1997-02-01). "'Sniffin' Sticks': Olfactory Performance Assessed by the Combined Testing of Odor Identification, Odor Discrimination and Olfactory Threshold". Chemical Senses. 22 (1): 39–52. doi: 10.1093/chemse/22.1.39 . ISSN 0379-864X. PMID 9056084.
- ↑ Atanasova, B (2008). "Olfaction: A potential cognitive marker of psychiatric disorder" (PDF). Neuroscience and Biobehavioral Reviews. 32 (7): 1315–1325. doi:10.1016/j.neubiorev.2008.05.003. PMID 18555528. S2CID 207088028.