
A toxin is a naturally occurring organic poison produced by metabolic activities of living cells or organisms. They occur especially as proteins, often conjugated. The term was first used by organic chemist Ludwig Brieger (1849–1919) and is derived from the word "toxic".

An algal bloom or algae bloom is a rapid increase or accumulation in the population of algae in freshwater or marine water systems. It is often recognized by the discoloration in the water from the algae's pigments. The term algae encompasses many types of aquatic photosynthetic organisms, both macroscopic multicellular organisms like seaweed and microscopic unicellular organisms like cyanobacteria. Algal bloom commonly refers to the rapid growth of microscopic unicellular algae, not macroscopic algae. An example of a macroscopic algal bloom is a kelp forest.
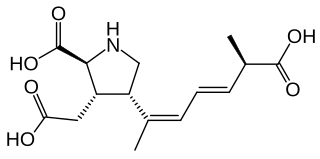
Domoic acid (DA) is a kainic acid-type neurotoxin that causes amnesic shellfish poisoning (ASP). It is produced by algae and accumulates in shellfish, sardines, and anchovies. When sea lions, otters, cetaceans, humans, and other predators eat contaminated animals, poisoning may result. Exposure to this compound affects the brain, causing seizures, and possibly death.

Saxitoxin (STX) is a potent neurotoxin and the best-known paralytic shellfish toxin (PST). Ingestion of saxitoxin by humans, usually by consumption of shellfish contaminated by toxic algal blooms, is responsible for the illness known as paralytic shellfish poisoning (PSP).
Okadaic acid, C44H68O13, is a toxin produced by several species of dinoflagellates, and is known to accumulate in both marine sponges and shellfish. One of the primary causes of diarrhetic shellfish poisoning, okadaic acid is a potent inhibitor of specific protein phosphatases and is known to have a variety of negative effects on cells. A polyketide, polyether derivative of a C38 fatty acid, okadaic acid and other members of its family have shined light upon many biological processes both with respect to dinoflagellete polyketide synthesis as well as the role of protein phosphatases in cell growth.
Paralytic shellfish poisoning (PSP) is one of the four recognized syndromes of shellfish poisoning, which share some common features and are primarily associated with bivalve mollusks. These shellfish are filter feeders and accumulate neurotoxins, chiefly saxitoxin, produced by microscopic algae, such as dinoflagellates, diatoms, and cyanobacteria. Dinoflagellates of the genus Alexandrium are the most numerous and widespread saxitoxin producers and are responsible for PSP blooms in subarctic, temperate, and tropical locations. The majority of toxic blooms have been caused by the morphospecies Alexandrium catenella, Alexandrium tamarense, Gonyaulax catenella and Alexandrium fundyense, which together comprise the A. tamarense species complex. In Asia, PSP is mostly associated with the occurrence of the species Pyrodinium bahamense.

Thin layers are concentrated aggregations of phytoplankton and zooplankton in coastal and offshore waters that are vertically compressed to thicknesses ranging from several centimeters up to a few meters and are horizontally extensive, sometimes for kilometers. Generally, thin layers have three basic criteria: 1) they must be horizontally and temporally persistent; 2) they must not exceed a critical threshold of vertical thickness; and 3) they must exceed a critical threshold of maximum concentration. The precise values for critical thresholds of thin layers has been debated for a long time due to the vast diversity of plankton, instrumentation, and environmental conditions. Thin layers have distinct biological, chemical, optical, and acoustical signatures which are difficult to measure with traditional sampling techniques such as nets and bottles. However, there has been a surge in studies of thin layers within the past two decades due to major advances in technology and instrumentation. Phytoplankton are often measured by optical instruments that can detect fluorescence such as LIDAR, and zooplankton are often measured by acoustic instruments that can detect acoustic backscattering such as ABS. These extraordinary concentrations of plankton have important implications for many aspects of marine ecology, as well as for ocean optics and acoustics. Zooplankton thin layers are often found slightly under phytoplankton layers because many feed on them. Thin layers occur in a wide variety of ocean environments, including estuaries, coastal shelves, fjords, bays, and the open ocean, and they are often associated with some form of vertical structure in the water column, such as pycnoclines, and in zones of reduced flow.

Karenia brevis is a microscopic, single-celled, photosynthetic organism in the genus Karenia. It is a marine dinoflagellate commonly found in the waters of the Gulf of Mexico. It is the organism responsible for the "Florida red tides" that affect the Gulf coasts of Florida and Texas in the U.S., and nearby coasts of Mexico. K. brevis has been known to travel great lengths around the Florida peninsula and as far north as the Carolinas.

Heterosigma akashiwo is a species of microscopic algae of the class Raphidophyceae. It is a swimming marine alga that episodically forms toxic surface aggregations known as harmful algal bloom. The species name akashiwo is from the Japanese for "red tide".
Neurotoxic shellfish poisoning (NSP) is caused by the consumption of brevetoxins, which are marine toxins produced by the dinoflagellate Karenia brevis. These toxins can produce a series of gastrointestinal and neurological effects. Outbreaks of NSP commonly take place following harmful algal bloom (HAB) events, commonly referred to as "Florida red tide". Algal blooms are a naturally-occurring phenomenon, however their frequency has been increasing in recent decades at least in-part due to human activities, climate changes, and the eutrophication of marine waters. HABs have been occurring for all of documented history, evidenced by the Native Americans' understanding of the dangers of shellfish consumption during periods of marine bioluminescence. Blooms have been noted to occur as far north as North Carolina and are commonly seen alongside the widespread death of fish and sea birds. In addition to the effects on human health, the economic impact of HAB-associated shellfish toxin outbreaks can have significant economic implications as well due to not only the associated healthcare costs, but the adverse impact on the commercial shellfish industry.

A harmful algal bloom (HAB), or excessive algae growth, is an algal bloom that causes negative impacts to other organisms by production of natural algae-produced toxins, mechanical damage to other organisms, or by other means. HABs are sometimes defined as only those algal blooms that produce toxins, and sometimes as any algal bloom that can result in severely lower oxygen levels in natural waters, killing organisms in marine or fresh waters. Blooms can last from a few days to many months. After the bloom dies, the microbes that decompose the dead algae use up more of the oxygen, generating a "dead zone" which can cause fish die-offs. When these zones cover a large area for an extended period of time, neither fish nor plants are able to survive. Harmful algal blooms in marine environments are often called "red tides".
Karenia is a genus that consists of unicellular, photosynthetic, planktonic organisms found in marine environments. The genus currently consists of 12 described species. They are best known for their dense toxic algal blooms and red tides that cause considerable ecological and economical damage; some Karenia species cause severe animal mortality. One species, Karenia brevis, is known to cause respiratory distress and neurotoxic shellfish poisoning (NSP) in humans.
Phycotoxins are complex allelopathic chemicals produced by eukaryotic and prokaryotic algal secondary metabolic pathways. More simply, these are toxic chemicals synthesized by photosynthetic organisms. These metabolites are not harmful to the producer but may be toxic to either one or many members of the marine food web. This page focuses on phycotoxins produced by marine microalgae; however, freshwater algae and macroalgae are known phycotoxin producers and may exhibit analogous ecological dynamics. In the pelagic marine food web, phytoplankton are subjected to grazing by macro- and micro-zooplankton as well as competition for nutrients with other phytoplankton species. Marine bacteria try to obtain a share of organic carbon by maintaining symbiotic, parasitic, commensal, or predatory interactions with phytoplankton. Other bacteria will degrade dead phytoplankton or consume organic carbon released by viral lysis. The production of toxins is one strategy that phytoplankton use to deal with this broad range of predators, competitors, and parasites. Smetacek suggested that "planktonic evolution is ruled by protection and not competition. The many shapes of plankton reflect defense responses to specific attack systems". Indeed, phytoplankton retain an abundance of mechanical and chemical defense mechanisms including cell walls, spines, chain/colony formation, and toxic chemical production. These morphological and physiological features have been cited as evidence for strong predatory pressure in the marine environment. However, the importance of competition is also demonstrated by the production of phycotoxins that negatively impact other phytoplankton species. Flagellates are the principle producers of phycotoxins; however, there are known toxigenic diatoms, cyanobacteria, prymnesiophytes, and raphidophytes. Because many of these allelochemicals are large and energetically expensive to produce, they are synthesized in small quantities. However, phycotoxins are known to accumulate in other organisms and can reach high concentrations during algal blooms. Additionally, as biologically active metabolites, phycotoxins may produce ecological effects at low concentrations. These effects may be subtle, but have the potential to impact the biogeographic distributions of phytoplankton and bloom dynamics.
Euglenophycin is an ichthyotoxic compound isolated from Euglena sanguinea, a protist of the genus Euglena. It exhibits anticancer and herbicidal activity in vitro.
Karenia mikimotoi is a dinoflagellate species from the genus Karenia. Its first appearance was in Japan in 1935 and since then, it has appeared in other parts of the world such as the east coast of the United States, Norway, and the English Channel.
Dinotoxins are a group of toxins which are produced by flagellate, aquatic, unicellular protists called dinoflagellates. Dinotoxin was coined by Hardy and Wallace in 2012 as a general term for the variety of toxins produced by dinoflagellates. Dinoflagellates are an enormous group of marine life, with much diversity. With great diversity comes many different toxins, however, there are a few toxins that multiple species have in common.

Antillatoxin (ATX) is a potent lipopeptide neurotoxin produced by the marine cyanobacterium Lyngbya majuscula. ATX activates voltage-gated sodium channels, which can cause cell depolarisation, NMDA-receptor overactivity, excess calcium influx and neuronal necrosis.

Dinoflagellates are eukaryotic plankton, existing in marine and freshwater environments. Previously, dinoflagellates had been grouped into two categories, phagotrophs and phototrophs. Mixotrophs, however include a combination of phagotrophy and phototrophy. Mixotrophic dinoflagellates are a sub-type of planktonic dinoflagellates and are part of the phylum Dinoflagellata. They are flagellated eukaryotes that combine photoautotrophy when light is available, and heterotrophy via phagocytosis. Dinoflagellates are one of the most diverse and numerous species of phytoplankton, second to diatoms.
Jan H. Landsberg is a biologist, researcher, and author. Her professional research interests in biology have particular focus on aquatic animal and environmental health.
Prymnesin-2 is an organic compound that is secreted by the haptophyte Prymnesium parvum. It belongs to the prymnesin family and has potent hemolytic and ichthyotoxic properties. In a purified form it appears as a pale yellow solid. P. parvum is responsible for red harmful algal blooms worldwide, causing massive fish killings. When these algal blooms occur, this compound poses a threat to the local fishing industry. This is especially true for brackish water, as the compound can reach critical concentrations more easily.




