Chromatin is a complex of DNA and protein found in eukaryotic cells. The primary function is to package long DNA molecules into more compact, denser structures. This prevents the strands from becoming tangled and also plays important roles in reinforcing the DNA during cell division, preventing DNA damage, and regulating gene expression and DNA replication. During mitosis and meiosis, chromatin facilitates proper segregation of the chromosomes in anaphase; the characteristic shapes of chromosomes visible during this stage are the result of DNA being coiled into highly condensed chromatin.

In biology, histones are highly basic proteins abundant in lysine and arginine residues that are found in eukaryotic cell nuclei. They act as spools around which DNA winds to create structural units called nucleosomes. Nucleosomes in turn are wrapped into 30-nanometer fibers that form tightly packed chromatin. Histones prevent DNA from becoming tangled and protect it from DNA damage. In addition, histones play important roles in gene regulation and DNA replication. Without histones, unwound DNA in chromosomes would be very long. For example, each human cell has about 1.8 meters of DNA if completely stretched out; however, when wound about histones, this length is reduced to about 90 micrometers (0.09 mm) of 30 nm diameter chromatin fibers.

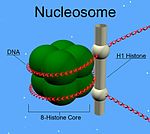
A nucleosome is the basic structural unit of DNA packaging in eukaryotes. The structure of a nucleosome consists of a segment of DNA wound around eight histone proteins and resembles thread wrapped around a spool. The nucleosome is the fundamental subunit of chromatin. Each nucleosome is composed of a little less than two turns of DNA wrapped around a set of eight proteins called histones, which are known as a histone octamer. Each histone octamer is composed of two copies each of the histone proteins H2A, H2B, H3, and H4.

In cell biology, the spindle apparatus refers to the cytoskeletal structure of eukaryotic cells that forms during cell division to separate sister chromatids between daughter cells. It is referred to as the mitotic spindle during mitosis, a process that produces genetically identical daughter cells, or the meiotic spindle during meiosis, a process that produces gametes with half the number of chromosomes of the parent cell.
Histone H2B is one of the 5 main histone proteins involved in the structure of chromatin in eukaryotic cells. Featuring a main globular domain and long N-terminal and C-terminal tails, H2B is involved with the structure of the nucleosomes.
The family of heterochromatin protein 1 (HP1) consists of highly conserved proteins, which have important functions in the cell nucleus. These functions include gene repression by heterochromatin formation, transcriptional activation, regulation of binding of cohesion complexes to centromeres, sequestration of genes to the nuclear periphery, transcriptional arrest, maintenance of heterochromatin integrity, gene repression at the single nucleosome level, gene repression by heterochromatization of euchromatin, and DNA repair. HP1 proteins are fundamental units of heterochromatin packaging that are enriched at the centromeres and telomeres of nearly all eukaryotic chromosomes with the notable exception of budding yeast, in which a yeast-specific silencing complex of SIR proteins serve a similar function. Members of the HP1 family are characterized by an N-terminal chromodomain and a C-terminal chromoshadow domain, separated by a hinge region. HP1 is also found at some euchromatic sites, where its binding can correlate with either gene repression or gene activation. HP1 was originally discovered by Tharappel C James and Sarah Elgin in 1986 as a factor in the phenomenon known as position effect variegation in Drosophila melanogaster.

The solenoid structure of chromatin is a model for the structure of the 30 nm fibre. It is a secondary chromatin structure which helps to package eukaryotic DNA into the nucleus.

Histone H3.1 is a protein that in humans is encoded by the HIST1H3B gene.

Core histone macro-H2A.1 is a protein that in humans is encoded by the H2AFY gene.

Non-histone chromosomal protein HMG-14 is a protein that in humans is encoded by the HMGN1 gene.

Histone H1.1 is a protein that in humans is encoded by the HIST1H1A gene.

Histone H3.1 is a protein in humans that is encoded by the H3C1 gene.
Histone H1.4 is a protein that in humans is encoded by the HIST1H1E gene.

Histone H3.1 is a protein that in humans is encoded by the HIST1H3F gene.
Histone H1.3 is a protein that in humans is encoded by the HIST1H1D gene.

Histone H1x is a protein that in humans is encoded by the H1FX gene.

Histone H2A.J is a protein that in humans is encoded by the H2AFJ gene.

Histone H2A-Bbd type 1 also known as H2A Barr body-deficient is a histone protein variant that in humans is encoded by the H2AFB1 gene.
Histone H2A-Bbd type 2/3 also known as H2A Barr body-deficient is a histone protein that in humans is encoded by the H2AFB2 gene.

In molecular biology, the linker histone H1 is a protein family forming a critical component of eukaryotic chromatin. H1 histones bind to the linker DNA exiting from the nucleosome core particle, while the core histones form the octamer core of the nucleosome around which the DNA is wrapped.