Artiodactyls are placental mammals belonging to the order Artiodactyla. Typically, they are ungulates which bear weight equally on two of their five toes: the third and fourth, often in the form of a hoof. The other three toes are either present, absent, vestigial, or pointing posteriorly. By contrast, most perissodactyls bear weight on an odd number of the five toes. Another difference between the two is that many artiodactyls digest plant cellulose in one or more stomach chambers rather than in their intestine as perissodactyls do. The advent of molecular biology, along with new fossil discoveries, found that cetaceans fall within this taxonomic branch, being most closely related to hippopotamuses. Some modern taxonomists thus apply the name Cetartiodactyla to this group, while others opt to include cetaceans within the existing name of Artiodactyla. Some researchers use "even-toed ungulates" to exclude cetaceans and only include terrestrial artiodactyls, making the term paraphyletic in nature.

Dietary fiber or roughage is the portion of plant-derived food that cannot be completely broken down by human digestive enzymes. Dietary fibers are diverse in chemical composition, and can be grouped generally by their solubility, viscosity, and fermentability, which affect how fibers are processed in the body. Dietary fiber has two main components: soluble fiber and insoluble fiber, which are components of plant-based foods, such as legumes, whole grains and cereals, vegetables, fruits, and nuts or seeds. A diet high in regular fiber consumption is generally associated with supporting health and lowering the risk of several diseases. Dietary fiber consists of non-starch polysaccharides and other plant components such as cellulose, resistant starch, resistant dextrins, inulin, lignins, chitins, pectins, beta-glucans, and oligosaccharides.
Digestion is the breakdown of large insoluble food compounds into small water-soluble components so that they can be absorbed into the blood plasma. In certain organisms, these smaller substances are absorbed through the small intestine into the blood stream. Digestion is a form of catabolism that is often divided into two processes based on how food is broken down: mechanical and chemical digestion. The term mechanical digestion refers to the physical breakdown of large pieces of food into smaller pieces which can subsequently be accessed by digestive enzymes. Mechanical digestion takes place in the mouth through mastication and in the small intestine through segmentation contractions. In chemical digestion, enzymes break down food into the small compounds that the body can use.

The Bovidae comprise the biological family of cloven-hoofed, ruminant mammals that includes cattle, yaks, bison, buffalo, antelopes, sheep and goats. A member of this family is called a bovid. With 143 extant species and 300 known extinct species, the family Bovidae consists of 11 major subfamilies and thirteen major tribes. The family evolved 20 million years ago, in the early Miocene.

Chewing or mastication is the process by which food is crushed and ground by the teeth. It is the first step in the process of digestion, allowing a greater surface area for digestive enzymes to break down the foods.
Cud is a portion of food that returns from a ruminant's stomach to the mouth to be chewed for the second time. More precisely, it is a bolus of semi-degraded food regurgitated from the reticulorumen of a ruminant. Cud is produced during the physical digestive process of rumination.
A monogastric organism has a simple single-chambered stomach. Examples of monogastric herbivores are horses and rabbits. Examples of monogastric omnivores include humans, pigs, hamsters and rats. Furthermore, there are monogastric carnivores such as cats. A monogastric organism is comparable to ruminant organisms, such as cattle, goats, or sheep. Herbivores with monogastric digestion can digest cellulose in their diets by way of symbiotic gut bacteria. However, their ability to extract energy from cellulose digestion is less efficient than in ruminants.
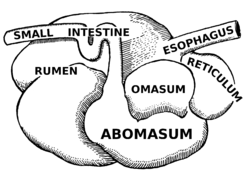
The rumen, also known as a paunch, is the largest stomach compartment in ruminants and the larger part of the reticulorumen, which is the first chamber in the alimentary canal of ruminant animals. The rumen's microbial favoring environment allows it to serve as the primary site for microbial fermentation of ingested feed. The smaller part of the reticulorumen is the reticulum, which is fully continuous with the rumen, but differs from it with regard to the texture of its lining.
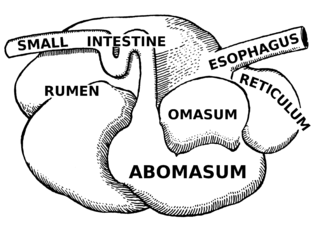
The omasum, also known as the bible, the fardel, the manyplies and the psalterium, is the third compartment of the stomach in ruminants. The omasum comes after the rumen and reticulum and before the abomasum. Different ruminants have different omasum structures and function based on the food that they eat and how they developed through evolution.

Enteric fermentation is a digestive process by which carbohydrates are broken down by microorganisms into simple molecules for absorption into the bloodstream of an animal. Because of human agricultural reliance in many parts of the world on animals which digest by enteric fermentation, it is the second largest anthropogenic factor for the increase in methane emissions directly after fossil fuel use.
The abomasum, also known as the maw, rennet-bag, or reed tripe, is the fourth and final stomach compartment in ruminants. It secretes rennet, which is used in cheese creation.
Cecotropes are a nutrient filled package created in the gastointestinal (GI) tract, expelled and eaten by rabbits and guinea pigs to get more nutrition out of their food. The first time through the GI tract, small particles of fiber are moved into the cecum, where microbes ferment them. This creates useable nutrients which are stored and expelled in cecotropes. The cecotropes are eaten and the nutrients are absorbed in the small intestine.

Pecora is an infraorder of even-toed hoofed mammals with ruminant digestion. Most members of Pecora have cranial appendages projecting from their frontal bones; only two extant genera lack them, Hydropotes and Moschus. The name "Pecora" comes from the Latin word pecus, which means "cattle". Although most pecorans have cranial appendages, only some of these are properly called "horns", and many scientists agree that these appendages did not arise from a common ancestor, but instead evolved independently on at least two occasions. Likewise, while Pecora as a group is supported by both molecular and morphological studies, morphological support for interrelationships between pecoran families is disputed.
Fibrobacter succinogenes is a cellulolytic bacterium species in the genus Fibrobacter. It is present in the rumen of cattle. F. succinogenes is a gram negative, rod-shaped, obligate anaerobe that is a major contributor to cellulose digestion. Since its discovery in the 1950s, it has been studied for its role in herbivore digestion and cellulose fermentation, which can be utilized in biofuel production.
Hindgut fermentation is a digestive process seen in monogastric herbivores. Cellulose is digested with the aid of symbiotic bacteria. The microbial fermentation occurs in the digestive organs that follow the small intestine: the large intestine and cecum. Examples of hindgut fermenters include proboscideans and large odd-toed ungulates such as horses and rhinos, as well as small animals such as rodents, rabbits and koalas. In contrast, foregut fermentation is the form of cellulose digestion seen in ruminants such as cattle which have a four-chambered stomach, as well as in sloths, macropodids, some monkeys, and one bird, the hoatzin.
Ruminal tympany, also known as ruminal bloat, is a disease of ruminant animals, characterized by an excessive volume of gas in the rumen. Ruminal tympany may be primary, known as frothy bloat, or secondary, known as free-gas bloat.
Methanogens are a group of microorganisms that produce methane as a byproduct of their metabolism. They play an important role in the digestive system of ruminants. The digestive tract of ruminants contains four major parts: rumen, reticulum, omasum and abomasum. The food with saliva first passes to the rumen for breaking into smaller particles and then moves to the reticulum, where the food is broken into further smaller particles. Any indigestible particles are sent back to the rumen for rechewing. The majority of anaerobic microbes assisting the cellulose breakdown occupy the rumen and initiate the fermentation process. The animal absorbs the fatty acids, vitamins and nutrient content on passing the partially digested food from the rumen to the omasum. This decreases the pH level and initiates the release of enzymes for further breakdown of the food which later passes to the abomasum to absorb remaining nutrients before excretion. This process takes about 9–12 hours.
Displaced abomasum in cattle occurs when the abomasum, also known as the true stomach, which typically resides on the floor of the abdomen, fills with gas and rises to the top of the abdomen, where it is said to be ‘displaced’. When the abomasum moves from its normal position it prevents the natural passage of gas and feed through the digestive system, creating a restriction. As cattle are ruminants, which have a 4 chambered stomach composed of a rumen, reticulum, omasum and abomasum. Ruminants require this specialized digestive system in order to properly process and break down their high fiber and cellulose rich diets. As this type of digestive system is quite complex it is at a greater risk for incidence. Due to the natural anatomy of cattle it is more common to have the abomasum displace to the left, known as a left-displaced abomasum, than to the right, right-displaced abomasum. When the abomasum becomes displaced there also becomes a chance of an abomasal volvulus, twist, developing. An abomasal volvulus occurs when the abomasum, which is already out of place, will rotate and cut off blood and nutrient supply to the abomasum. Cattle which develop an abomasal twist require immediate vet attention to regain blood supply and food passage through the digestive system or the abomasum will begin to shut down due to lack of blood supply and toxicity development.

The Jarman–Bell principle is a concept in ecology that the food quality of a herbivore's intake decreases as the size of the herbivore increases, but the amount of such food increases to counteract the low quality foods. It operates by observing the allometric properties of herbivores. The principle was coined by P.J Jarman (1968.) and R.H.V Bell (1971).

Anaeromyces robustus is a fungal microorganism that lives in the gut rumen of many ruminant herbivores such as cows and sheep. Previously thought to be protozoa from their flagellated zoospores, they are biomass degraders, breaking down carbohydrates and plant materials from the food the animal ingests. This fungus, therefore, is anaerobic and lives without oxygen. Gut fungi are dramatically outnumbered by other organisms in the microbiome; they are members of the gut microbiome in ruminants and hindgut fermenters and play a key role in digestion.